1. L'attention
L'étude de l'attention s'est beaucoup développée depuis une cinquantaine d'années et a permis de mettre en évidence l'existence de différents processus, ou mécanismes, attentionnels. L'attention ne serait pas constituée d'un processus psychologique unique mais d'un ensemble de processus qui seraient en réseau les uns avec les autres. Cohen (1993) par exemple distingue quatre types de processus: la sélection de l'information, les ressources attentionnelles, le contrôle de la réponse et de l'activité, et l'attention soutenue. Les études effectuées dans le domaine de l'attention ont essentiellement cherché à préciser les caractéristiques de ces processus en spécifiant leur importance, leur rôle et leur influence sur les processus cognitifs en général (perception, langage, mémoire, etc.). Beaucoup d'études ont également porté sur les localisations cérébrales des zones sous-jacentes à ces processus. L'existence d'un processus attentionnel d'ordre spatial a été exploré dans de nombreuses études chez le sujet sain (par exemple Posner et al., 1980; Treisman, 1986). Un grand nombre d'indices confirment l'existence d'un tel processus attentionnel qui serait spécialisé dans la détection et la sélection de l'information (attention sélective) en fonction de la position spatiale des objets et des événements, il contrôlerait également le flux d'informations entre les systèmes perceptifs et les systèmes 'cognitifs de plus haut niveau'. Les travaux que nous présentons avaient pour but de spécifier les bases neurales de ce type de processus attentionnel spatial et de comparer ce processus dans la modalité auditive et la modalité visuelle. L'étude de patients héminégligents présentant des lésions cérébrales localisées dans l'hémisphère droit avaient pour objectif de déterminer l'importance de certaines régions cérébrales dans le fonctionnement des processus attentionnels. Nous présentons ci-dessous différents travaux ayant essentiellement étudié ces types de processus dans des études comportementales ou d'imagerie cérébrale fonctionnelle.
Les études comportementales portent généralement sur certains processus attentionnels tels que la détection d'un stimulus nouveau ou incongru, l'orientation spatiale de l'attention ou encore l'attention sélective. Ces travaux ont permis de mettre en évidence la présence de tels processus et de déterminer leur influence respective sur le comportement (amélioration ou détérioration des performances par exemple). Les études d'imagerie cérébrale ont permis de préciser les zones du cerveau sous-jacentes à certains de ces processus. Un grand nombre de travaux ayant été effectué dans ce domaine, nous avons sélectionné plusieurs études portant plus spécifiquement sur les processus attentionnels que nous avons étudié. Nous avons ensuite abordé le domaine de l'intermodalité dans l'attention afin de préciser les similitudes, les différences et les connexions qui pourraient exister entre les processus attentionnels auditifs et les processus attentionnels visuels. Finalement, quelques modèles portant sur l'organisation des processus attentionnels sont abordés. Nous n'avons pas présenté tous les modèles attentionnels existants dans la littérature, nous nous sommes concentrés sur les modèles qui nous ont semblé être les plus proches de notre problématique.
1.1. Les bases neurales des processus attentionnels
1.1.1. Etudes comportementales
Les travaux ayant utilisé des techniques comportementales dans l'étude des processus attentionnels sont nombreux. Ces techniques portent essentiellement le temps de réaction et le nombre de réponses (réponses correctes ou fausses, et absences de réponses). Nous avons choisi de présenter trois groupes d'études ayant porté sur différents processus attentionnels correspondant à ceux que nous avons étudiés dans les travaux présentés dans les chapitres 6 à 9. Nous présentons ci-dessous les travaux portant sur l'étude des processus d'orientation visuo-spatiale de l'attention effectués par Posner et ses collaborateurs (pour une revue voir Posner et Raichle, 1994). Les études de Schneider et Shiffrin (1977) sont ensuite abordées. Elles ont permis de dissocier les processus automatiques des processus contrôlés. Après la description de ces recherches effectuées dans la modalité visuelle, nous décrivons des études dans la modalité auditive portant sur le paradigme d'écoute dichotique. Ces derniers travaux ont suggéré la présence d'effets facilitant (ou améliorant) le traitement de stimuli faisant l'objet d'une focalisation de l'attention.
1.1.1.1. L'orientation visuo-spatiale de l'attention
Les travaux de Posner et de ses collaborateurs ont principalement porté sur l'attention visuo-spatiale chez les sujets normaux et chez des patients cérébro-lésés. Dans une expérience type, le sujet fixe un point central sur un écran et des stimuli (cibles) sont présentés à gauche ou à droite du point de fixation (Fig. 1). Les sujets doivent appuyer sur un bouton dès qu'ils perçoivent un stimulus quel que soit l'emplacement de ce stimulus (cible). Un stimulus précède l'apparition de ces cibles, il s'agit des stimuli 'indices'. Ces indices correspondent par exemple à l'apparition d'un carré jaune lumineux où la cible a le plus de chance d'apparaître (à droite ou à gauche). Dans certains cas, un signe '+' peut également apparaître, il indique que la cible a autant de chance d'apparaître à gauche qu'à droite.
Pour résumer les principaux résultats de ces études, ces expériences montrent essentiellement que les temps de réaction des sujets sont influencés par les informations déduites des indices. Ces auteurs ont observé globalement que lorsque l'indice était un signe '+' (condition 'neutre'), le temps de réaction était d'environ 250-300 ms. Lorsque l'indice indiquait correctement le côté d'apparition de la cible (condition 'valide'), les temps de réaction étaient environ 30 ms plus courts que dans la condition 'neutre' et lorsque l'indice n'indiquait pas le bon côté (condition 'non valide'), les temps de réaction étaient environ 30 ms plus longs que dans la condition 'neutre'. L'effet de l'indice peut donc être soit un bénéfice (diminution du temps de réaction), soit un coût (augmentation du temps de réaction) selon sa validité.

Posner et ses collaborateurs (Posner et Raichle, 1994) ont développé un modèle décrivant le décours des opérations mentales qui seraient impliquées dans leur tâche (Fig. 2). Ils ont proposé également des localisations cérébrales (Fig. 3) pour certaines de ces opérations. Elles se situeraient dans des zones considérées actuellement comme étant impliquées dans les processus attentionnels (voir les études en imagerie fonctionnelle ci-dessous).
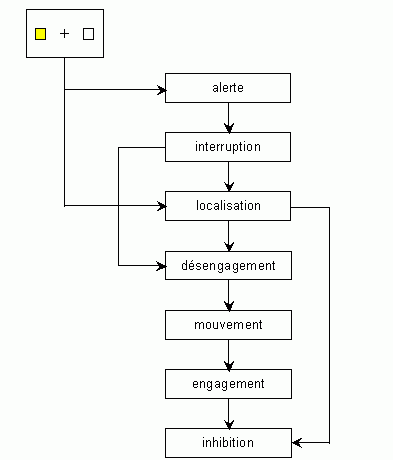
*L'indice capte et interrompt l'attention du sujet ('alerte' et 'interruption') qui, après avoir localisé la cible ('localisation') désengage son attention de l'emplacement précédant ('désengagement') pour l'amener ('mouvement') vers le nouvel emplacement. L'attention s'engage alors sur ce nouvel emplacement ('engagement') et l'ancien emplacement est alors inhibé ('inhibition'), adapté de Posner et Raichle, 1994.
L'ensemble des études effectuées par ces auteurs a mis en évidence certaines propriétés des processus d'orientation visuo-spatiale de l'attention. Par exemple, lorsque l'attention quitte un endroit pour revenir sur le point de fixation, le temps de réaction est plus lent lorsque la cible apparaît effectivement sur cet emplacement que lorsqu'elle apparaît dans un autre emplacement. Cet effet est généralement appelé 'inhibition de retour', il correspond à une difficulté à réorienter son attention dans une direction qui vient d'être attendue. Selon les auteurs, deux processus d'orientation de l'attention existeraient: un processus attentionnel exogène qui serait activé par l'apparition des indices, ce processus serait spontané, réflexe. L'autre processus serait de nature endogène, il serait mis en jeu lors de la condition 'neutre' dans laquelle le sujet porte volontairement son attention d'un côté ou de l'autre. Ce processus donnerait donc lieu à une orientation volontaire et contrôlée de l'attention. Les études de Posner ont également montré que l'orientation de l'attention peut avoir lieu sans aucun mouvement du regard en direction de la zone sur laquelle l'attention se porte. Dans les tâches que nous avons utilisées, les sujets orientaient leur attention vers la droite ou la gauche selon la consigne, en gardant toujours le regard fixé devant eux. Nos travaux ont donc mis en jeu, notamment, une orientation endogène de l'attention selon la terminologie de Posner. Une telle dissociation entre deux processus est également proposée par Schneider et Shiffrin (1977): ces auteurs distinguent d'une part des processus automatiques et d'autre part des processus contrôlés (ou volontaires).

1.1.1.2. Les processus attentionnels automatiques et contrôlés
Dans leurs premières études portant sur les processus automatiques et contrôlés du traitement de l'information, Schneider et Shiffrin (Schneider et Shiffrin, 1977; Shiffrin et Schneider, 1977) ont développé un paradigme mettant en jeu des processus de recherche et de détection de cibles visuelles parmi des distracteurs. Ce paradigme était constitué de présentations successives de planches contenant aux quatre coins différents items: des chiffres, des lettres ou des configurations de points. Certaines planches pouvaient être complètement blanches. Une première planche indiquait au sujet la cible à détecter dans les planches suivantes. La cible pouvait être constituée d'un seul item (le chiffre '1' par exemple) ou de plusieurs items (les lettres 'A' et 'D' par exemple). Les planches présentées successivement pouvaient contenir ou non cette cible. Les cibles étaient déterminées pour chaque essai (un essai correspond à la présentation d'une planche). Le sujet devait simplement appuyer sur un 'bouton-réponse' dès qu'il percevait la cible de l'essai en question. Plusieurs variations expérimentales ont été étudiées: le nombre de cibles à détecter, le temps de présentation des planches, le nombre d'items sur une planche, la nature des règles déterminant si un item donné est une cible ou un distracteur. Cette dernière manipulation expérimentale s'est avérée être la plus importante. Dans certaines séquences d'essais, un item peut être considéré comme une cible à un essai et comme un distracteur à l'essai suivant. Les cibles sont différentes dans chaque essai. Dans d'autres séries d'essais, une cible n'apparaît jamais parmi les distracteurs et vice-versa, un ensemble de cibles est déterminé et ne change pas à travers les essais de ces séquences.
Plusieurs études ont été effectuées avec ce paradigme. Elles ont mis en évidence, principalement, que le type de séquences de stimulation (changement de cibles à chaque essai) et la construction des stimuli (nature des stimuli cibles similaire ou différente de celle des stimuli distracteurs entre les essais) sont les variables influençant le plus le temps de réaction et le nombre de détections correctes. Les effets observés témoignent de la participation de deux processus différents dans l'exécution de la tâche: un processus attentionnel automatique et un processus attentionnel contrôlé. Les principales caractéristiques de ces deux processus peuvent être résumées ainsi: les processus automatiques sont rapides, parallèles, ne demandent pas l'intervention de la conscience ni une grande charge attentionnelle. En revanche, les processus contrôlés sont lents, sériels, nécessitent la participation de la conscience et demandent une charge attentionnelle plus importante. Ces caractéristiques sont présentées plus en détail dans le chapitre 1.3. (Tabl. 1). Elles se rapprochent beaucoup de celles suggérées par Posner pour les processus exogènes et endogènes abordés ci-dessus (Posner et Raichle, 1994 notamment). Les études de Posner d'une part et de Schneider et Shiffrin d'autre part, mettent donc en évidence la présence de processus attentionnels distincts qui ont été à la base de modèles attentionnels développés par ces auteurs (présentés dans le chapitre 1.3.). Après avoir abordé deux groupes d'études portant sur l'attention visuelle, nous présentons ci-dessous des travaux étudiant l'attention auditive.
1.1.1.3. L'attention sélective dans l'écoute dichotique
L'étude de l'attention sélective a souvent été effectuée à l'aide d'expériences mettant en jeu simultanément plusieurs sources d'informations. Dans le domaine de l'attention sélective auditive, beaucoup d'études ont porté sur le paradigme de l'écoute dichotique développé par Cherry (1953). Ce paradigme consiste en une présentation de messages auditifs différents délivrés simultanément à chaque oreille. L'utilisation d'un casque d'écouteurs permet de s'assurer que le message délivré dans l'oreille droite ne parvient pas à l'oreille gauche et vice-versa. Les tâches habituellement effectuées dans ce paradigme demandent généralement au sujet de focaliser son attention sur une oreille, il se trouve alors dans une situation appelée 'attention focalisée' ou 'attention sélective'. Dans un autre type de tâche, le sujet doit répartir son attention sur les deux oreilles, cette situation est généralement appelée 'attention divisée'. Ce paradigme a permis de mettre en évidence les principaux effets liés à la sélection d'information et à la focalisation de l'attention auditive.
Geffen et ses collaborateurs (Geffen et Sexton, 1978; Geffen et Traub, 1979; Geffen et Whale, 1979) ont comparé les performances des sujets dans une tâche d'attention divisée à celles observées dans une tâche d'attention sélective. Les sujets devaient détecter des cibles (certains mots) présentées soit dans l'oreille droite soit dans l'oreille gauche parmi un ensemble de mots présentés dichotiquement. Lorsque les sujets sont en situation d'attention focalisée, la détection des cibles dans l'oreille attentive est nettement meilleure que celle observée dans les situations d'attention divisée (environ 65% par rapport à 45%).De plus, les performances de détection de cibles dans l'oreille inattentive lors de la situation d'attention focalisée est très faible (environ 10% de cibles détectées). Les principales conséquences de la sélection de l'information et de la focalisation de l'attention sont donc:
- une amélioration des performances en facilitant le traitement de l'information cible (stimulus cible ou oreille attentive),
- une altération des performances concernant les distracteurs (stimulus à ignorer ou oreille inattentive).
1.1.1.4. Conclusion
L'ensemble des travaux présenté ci-dessus a permis de mettre en évidence la présence de plusieurs processus attentionnels et de préciser leurs influences respectives sur les processus cognitifs. D'une façon générale, les effets de l'attention sur le comportement pourraient se résumer ainsi: l'attention renforce la détection et optimise le temps de réaction:
- les études de Posner portant sur l'orientation visuo-spatiale de l'attention ont mis en évidence des effets de bénéfices et de coûts lors de la présentation d'indices. Ces résultats suggèrent la présence d'un processus attentionnel endogène, ou volontaire, et d'un processus attentionnel exogène, ou automatique,
- les travaux de Schneider et Shiffrin utilisant des tâches de recherche et de détection de cibles ont également mis en évidence la présence d'un processus attentionnel automatique et d'un processus attentionnel contrôlé,
- les expériences utilisant le protocole d'écoute dichotique ont révélé des effets d'amélioration et de détérioration des performances de l'attention suggérant la présence de processus facilitant ou empêchant le traitement des stimuli.
Les mécanismes que nous avons étudiés dans ce travail correspondent aux processus attentionnels automatiques et contrôlés observés dans les deux premiers groupes d'études. Nous présentons ci-dessous les travaux effectués en imagerie cérébrale qui ont fourni des informations complémentaires sur ces processus d'une part et qui ont permis de préciser les zones cérébrales sous-jacentes à ces processus d'autre part.
1.1.2. Etudes en imagerie fonctionnelle
1.1.2.1. Les techniques d'imagerie fonctionnelle
La tomographie par émissions de positons est une technique qui se base sur la détection de signaux radioactifs (marqueurs) dans le sang (Raichle et al., 1983). La procédure de base consiste à injecter une solution radioactive dans la circulation sanguine. Une zone active dans le cerveau nécessite une consommation plus élevée d'oxygène qui est délivré par la microcirculation sanguine. L'isotope radioactif contenu dans la solution injectée est transporté dans les zones cérébrales activées qui présentent alors une grande quantité de cet isotope. Sa désintégration produit des positons qui aboutissent à l'émission de rayons gamma. Ces rayons sont détectales et quantifiables à l'extérieur du cerveau par l'intermédiaire d'une caméra à positons. A partir des variations de concentration de cet isotope en fonction de l'activité plus ou moins importante des zones cérébrales, cette technique permet de visualiser les zones du cerveau impliquées dans une tâche perceptive, sensori-motrice ou cognitive. Cette technique présente l'avantage d'avoir une résolution spatiale élevée, de l'ordre du millimètre, mais elle possède deux désavantages importants: une résolution temporelle faible et l'injection d'une solution radioactive (Dehaene, 1997).
La technique de l'IRMf permet également de visualiser les zones cérébrales actives lors d'une tâche mais elle se base sur des modulations du débit sanguin et de l'oxygénation du cerveau (Ogawa et al., 1992). Cependant, cette technique ne nécessite pas forcément l'injection d'un marqueur: elle utilise les effets magnétiques du sang dans un milieu présentant un champ magnétique spécifique. La méthode 'BOLD' (Blood Oxygen Level Dependent contrast) est actuellement beaucoup utilisée. Selon cette méthode, la déoxyhémoglobine (correspondant globalement à une molécule d'hémoglobine ne possédant plus de molécule d'oxygène) présente la propriété de pouvoir s'aimanter dans un champ magnétique contrairement aux molécules d'oxyhémoglobine. Lors d'une activation, il se produit une importante augmentation du débit sanguin local saturé en oxygène. Cette augmentation dépasse de beaucoup l'augmentation légère de la consommation en oxygène de la région activée. La différence entre ces deux augmentations conduit à une diminution nette de la quantité de déoxyhémoglobine dans la région activée. Cette diminution entraîne une petite remontée du signal magnétique captée par le système d'IRM (Dehaene, 1997).
Un des avantages de l'IRMf est sa grande précision spatiale (qui tend actuellement à être inférieure à un millimètre). De plus, sa résolution temporelle peu élevée au début de l'utilisation de cette technique, s'améliore de plus en plus (environ de l'ordre d'une seconde pour le moment). Cet avantage, additionné au fait de ne pas avoir à effectuer d'injection d'un traceur, a conduit beaucoup de chercheurs à utiliser principalement cette technique depuis quelques années. Cependant, ces deux techniques (TEP et IRMf) ne reflètent que des changements indirects d'activité. En effet, elles se basent sur le débit sanguin et les changements d'oxygénation qui restent lents comparés au fonctionnement des neurones. En revanche, deux autres techniques reflètent directement l'activité des neurones, il s'agit de l'électroencéphalographie et de la magnétoencéphalographie. A l'inverse des techniques TEP ou IRMf, l'électro- et la magnétoencéphalographie présentent l'avantage important d'avoir une très bonne résolution temporelle (de l'ordre de la milliseconde). Nous présentons ci-dessous certaines études ayant utilisé la TEP ou l'IRMf dans l'étude des processus attentionnels. Parmi le grand nombre d'études dans ce domaine, nous avons sélectionné celles qui nous ont paru les plus importantes dans le contexte de nos travaux.
1.1.2.2. Le rôle du lobe pariétal dans l'orientation spatiale
Beaucoup d'études ont montré en TEP l'activation du lobe pariétal dans différentes tâches. Plusieurs régions au niveau du cortex pariétal supérieur seraient actives par exemple lors d'un traitement spatial (Haxby et al., 1991), lors d'une sélection spatiale (Corbetta et al., 1993), dans une tâche de vigilance visuelle (Pardo et al., 1991). Au niveau du cortex pariétal inférieur, différentes régions seraient activées lorsque l'attention porte sur la vitesse d'un pattern visuel (Corbetta et al., 1990). Il semble donc que le lobe pariétal soit impliqué dans des processus d'attention visuelle.
Dans une tâche impliquant plus précisément une orientation spatiale de l'attention visuelle, Corbetta et al. (1993) ont étudié les variations d'activités cérébrales en fonction du type d'attention mise en jeu (attention endogène ou exogène) et en fonction du côté sur lequel l'attention se porte. Ces auteurs ont observé une augmentation d'activité dans le lobe pariétal supérieur et dans le lobe frontal supérieur. L'augmentation au niveau du lobe pariétal supérieur a été observée dans les situations de changements endogènes (volontaires) ou exogènes (automatiques) d'orientation de l'attention. Ils ont également mis en évidence des différences d'activités au niveau pariétal lorsque l'attention devait se porter sur gauche ou sur la droite. En effet, une augmentation d'activité au niveau du lobe pariétal supérieur droit est observée lorsque les sujets portent leur attention sur la gauche mais quand ils portent leur attention sur la droite, on observe une augmentation d'activité dans les lobes pariétaux supérieurs gauche et droit. De plus, les zones activées au niveau du lobe pariétal supérieur droit étaient distinctes selon le côté sur lequel l'attention se portait (Fig. 4).
Selon les auteurs, les processus qui se situeraient au niveau du lobe pariétal supérieur permettraient un changement d'orientation spatiale. Une activité dans cette région a été également observée durant des saccades oculaires vers le champ visuel périphérique, déclenchées volontairement ou par l'apparition d'un stimulus (Anderson, 1994; Petersen et al., 1994). Hunton et al. (1995) ont aussi observé une augmentation de l'activité dans cette région lors de la recherche visuelle de cibles définies selon leur couleur ou leur orientation. L'ensemble de ces résultats met en évidence que l'activité pariétale est présente lorsque la tâche demande un changement de l'orientation de l'attention. L'asymétrie que Corbetta et al. (1993) ont observée en fonction du côté de présentation des cibles suggère que la capacité de changer d'orientation attentionnel et de pouvoir attendre et détecter ainsi un stimulus visuel à un endroit donné implique donc principalement le cortex pariétal (supérieur) de l'hémisphère droit.
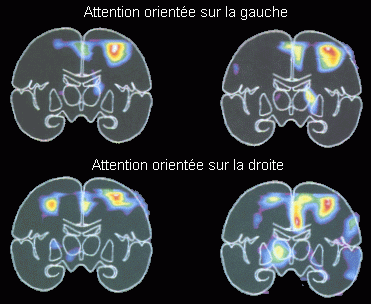
*lors de l'orientation de l'attention vers la gauche (en haut) ou vers la droite (en bas). Les coupes de gauche sont légèrement plus antérieures (24 mm) que les coupes de droite (34 mm). Lorsque l'attention est dirigée vers la gauche, une augmentation du débit apparaît au niveau du lobe pariétal supérieur droit. En revanche, lorsque l'attention est dirigée vers la droite, une augmentation apparaît dans les lobes pariétaux supérieurs droit et gauche
De plus, dans une étude utilisant l'IRMf, Corbetta et al. (2000) ont montré l'implication d'autres régions lors de tâches d'attention sélective. En effet, ces auteurs ont observé une première activation au niveau du sillon intrapariétal quand l'attention porte sur un endroit donné avant la présentation de la cible. Une augmentation d'activité apparaît ensuite au niveau de la jonction temporo-pariétale de l'hémisphère droit quand la cible est détectée, en particulier lorsqu'elle ne se situe pas à l'endroit attendu. Ces résultats mettent en évidence que les processus attentionnels d'attente de la cible (première activation) et les processus de détection de cette cible (deuxième activation) mettent en jeu des zones différentes.
Les travaux récents de Corbetta et de ses collaborateurs (2000) confirment donc les observations constatées dans les études présentées ci-dessus, mais ils précisent également que le lobe pariétal peut être impliqué non seulement dans un processus d'attention sélective (orientation) mais également dans un processus de détection de cibles (au niveau de la jonction temporo-pariétale). Il est donc nécessaire d'approfondir le rôle de régions différentes dans les processus attentionnels (détection, orientation,...). Cependant, l'ensemble des études présentées dans ce chapitre s'accorde sur l'implication du lobe pariétal lors de tâche d'attention spatiale visuelle. Cette région serait plus précisément responsable du changement d'orientation de l'attention.
1.1.2.3. Le gyrus cingulaire et les régions frontales dans les processus attentionnels
Dans leurs études sur l'attention visuelle, Corbetta et al. (1993) ont également observé une augmentation d'activité dans le cortex cingulaire antérieur. Un résultat similaire a été observé dans d'autres travaux, dont ceux utilisant des paradigmes d'interférence (Pardo et al., 1990). En effet, les travaux utilisant des paradigmes d'interférence, dont celui portant sur l'effet Stroop (Stroop, 1935) ont été également utilisés dans le but d'analyser les processus d'orientation de l'attention. Cet effet est observé lorsque le sujet doit dénommer la couleur de l'encre de mots désignant des couleurs alors que la couleur de l'encre et la couleur que le mot désigne ne sont pas pareilles (par exemple le sujet doit dire 'rouge' pour le mot 'bleu' écrit à l'encre rouge). Le temps de dénomination de la couleur de l'encre du mot est plus long que le temps de dénomination de la couleur de simples carrés colorés. Cette différence de temps provient de l'interférence du sens du mot écrit décrivant une couleur (qu'il s'agit donc d'inhiber) sur la dénomination de la couleur de l'encre elle-même. Pardo et al. (1990) ont montré en TEP, dans une telle tâche de dénomination de la couleur de l'encre du mot, une activité élevée dans la région du cortex cingulaire antérieur. Ces résultats suggèrent que cette zone soit sous-jacente à l'activité de sélection visuelle permettant une inhibition de la réponse automatique du sens du mot écrit. De plus, selon Corbetta et al. (1991), le gyrus cingulaire antérieur serait impliqué dans un système de sélection d'une réponse lors de processus attentionnels contrôlés.
D'une façon générale, l'attention auditive a été nettement moins étudiée que l'attention visuelle, notamment dans les études en imagerie cérébrale. Tzourio et al. (1997) rappellent d'ailleurs que la plupart des études ayant porté sur l'attention auditive utilisaient la technique des potentiels évoqués (voir chapitre 1.1.3.) Dans une étude en TEP portant sur l'attention sélective auditive, ces auteurs ont mis en évidence des augmentations d'activité au niveau du cortex cingulaire antérieur et des régions préfrontales droites. L'activité observée au niveau du gyrus cingulaire antérieur dans cette étude correspondrait à celle observée par Pardo et al. (1990) dans les tâches visuelles utilisant le paradigme de Stroop. Les résultats observés par Tzourio, confirmeraient donc l'implication du gyrus cingulaire antérieur dans les processus de sélection d'une réponse.
Dans les études mettant en évidence des activités au niveau du gyrus cingulaire antérieur, le cortex préfrontal présentait également une augmentation d'activité (notamment Corbetta et al., 1991; 1993; Pardo et al., 1990; 1991; Tzourio et al., 1997). Dans une tâche de vigilance concernant les modalités visuelle et somesthésique, Pardo et al. (1991) ont mis en évidence une implication de la région préfrontale droite (et du cortex supérieur pariétal droit également). Les activités observées au niveau du lobe frontal supérieur refléteraient un processus d'orientation active de l'attention.
1.1.2.4. Conclusion
Les différentes études en imagerie fonctionnelle mettent en évidence l'implication de plusieurs zones cérébrales dans les mécanismes attentionnels. En effet, des augmentations d'activité dans 4 régions cérébrales sont souvent observées dans toutes les études en TEP et en IRMf que nous avons présentées.
- le lobe pariétal postérieur (droit principalement)
- le cortex frontal ou préfrontal (droit principalement)
- le cortex cingulaire antérieur
- le colliculus supérieur
Les études électrophysiologiques que nous allons aborder apportent des informations complémentaires sur les processus attentionnels. En effet, les techniques TEP et IRMf fournissent des informations précises sur l'emplacement des zones cérébrales activées mais le décours temporel de ces activations ne peut pas être observé en détail par ces études. En revanche, la technique des potentiels évoqués, reflétant l'activité électrique des neurones, permet de rendre compte des activations cérébrales successives (mais sans grande précision spatiale).
1.1.3. Etudes électrophysiologiques
La technique des potentiels évoqués est une méthode particulièrement bien adaptée à l'étude du décours temporel des processus cognitifs. Elle permet de rendre compte, presque en temps réel, des activités cérébrales successives sous-jacentes à ces processus. La présence de composantes (pics d'amplitude) sur les potentiels évoqués témoignent de l'activité électrique d'une ou de plusieurs zones du cerveau. Les techniques d'acquisition et d'analyse des potentiels évoqués sont exposées en détail dans le chapitre 3. Nous avons indiqué cependant brièvement ci-dessous les trois types d'analyses utilisées dans les études décrites dans le présent chapitre. Ces analyses généralement portent sur:
- les variations d'amplitude et de latence des composantes (ou ondes) successives sur les courbes de potentiels (représentant l'amplitude en fonction du temps),
- la topographie de ces composantes sur le scalp à l'aide de cartes de potentiel représentant les valeurs des potentiels sur le scalp,
- la reconstruction des générateurs cérébraux sous-jacents aux composantes observées sur le scalp à partir de modèles physico-mathématiques de l'activité et de la propagation des signaux électriques dans le cerveau.
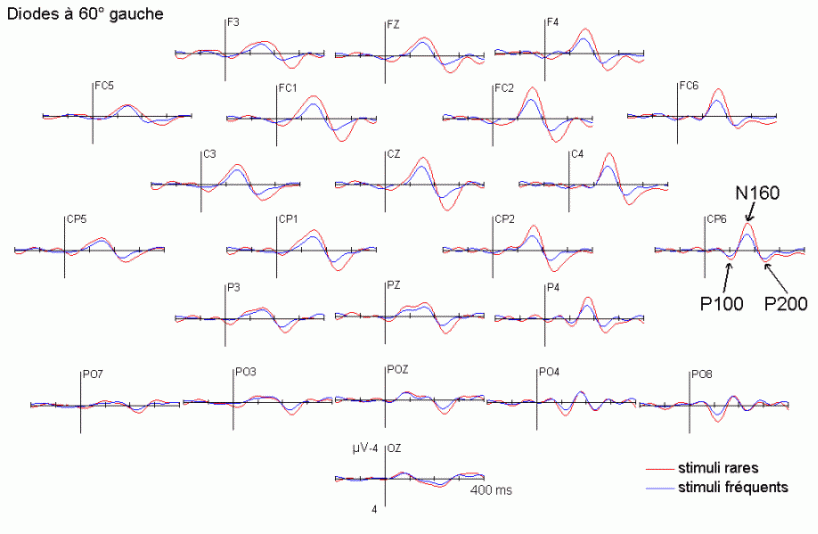
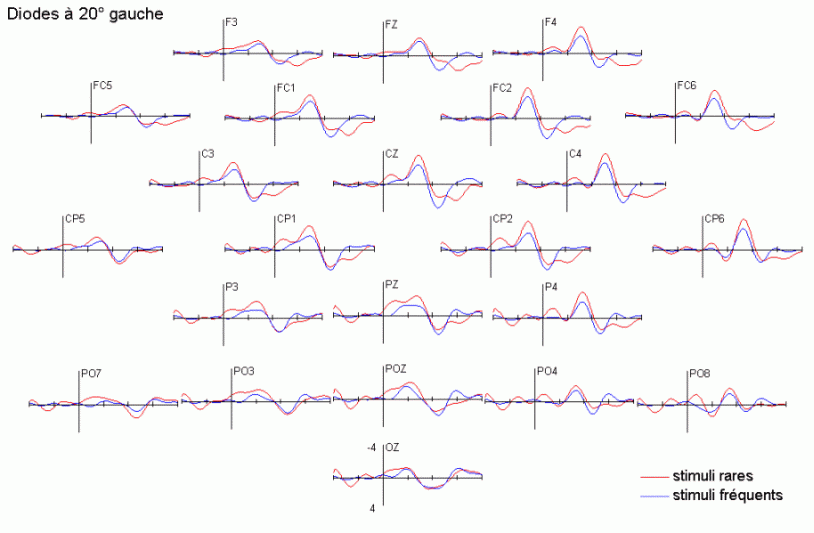
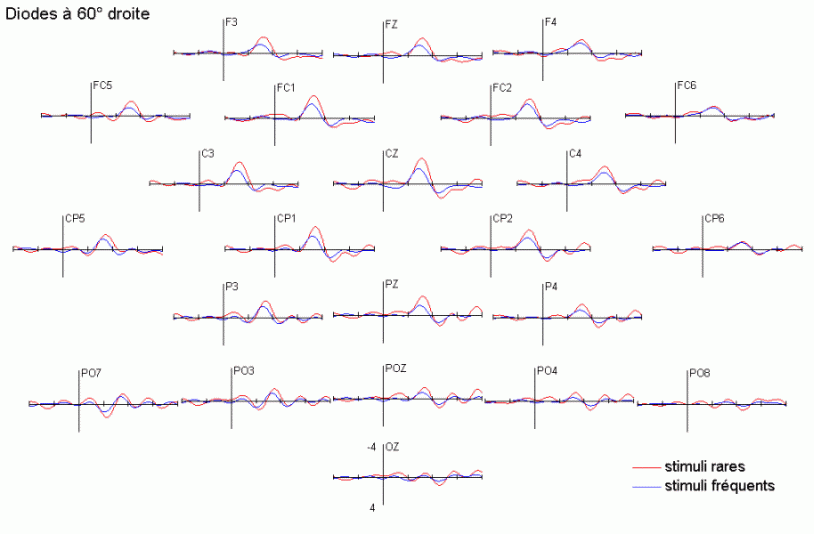
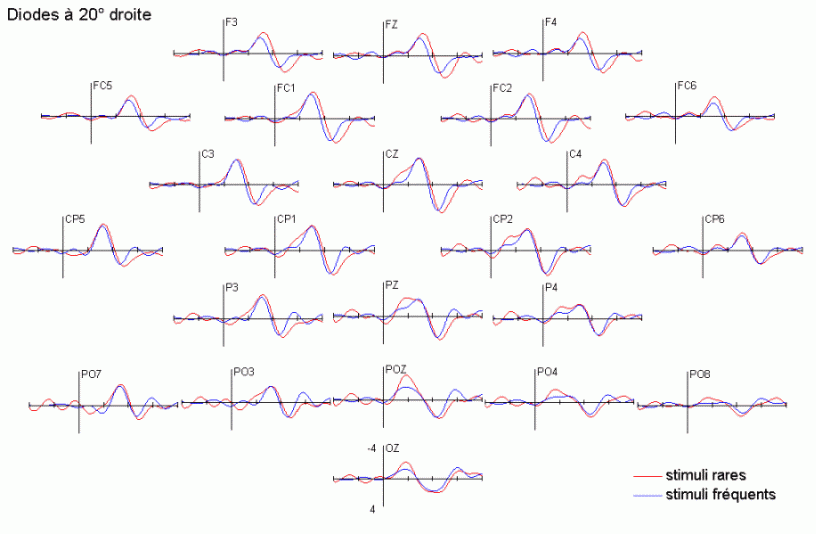
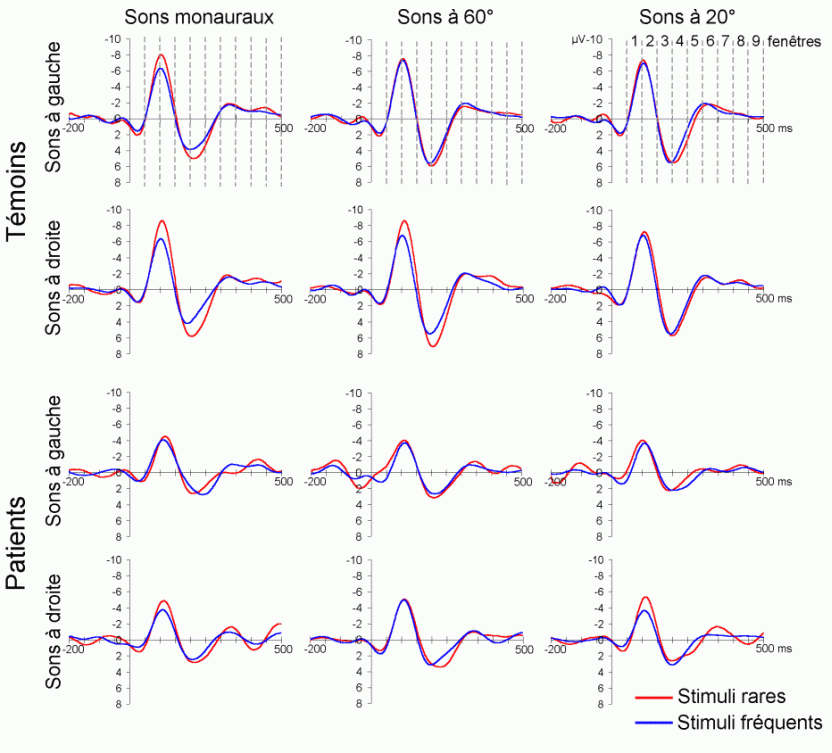
Dans le domaine des potentiels évoqués, l'attention a été essentiellement étudiée à l'aide des paradigmes de type 'oddball' ou d'attention sélective (Ritter et Vaughan, 1969). En audition comme en vision, ces paradigmes sont composés de séquences contenant des stimuli rares (ou déviants) ayant une probabilité d'apparition peu élevée (par exemple 20%) et des stimuli fréquents (ou standards), ayant une probabilité d'apparition nettement plus forte (par exemple 80%). Les stimuli rares peuvent avoir le statut de cibles à détecter le mieux et le plus rapidement possible. Ils peuvent également être présentés (alternativement avec les stimuli fréquents) sans qu'aucune tâche ne soit demandée au sujet. La majorité des études que nous présentons ci-dessous ont utilisé ce type de paradigme. Elles ont porté principalement sur les effets liés à l'attention automatique, essentiellement dans la modalité auditive, et sur l'attention sélective dans les modalités auditive et visuelle. Les travaux que nous avons exposés ci-dessous ont été regroupés en fonction des mécanismes qu'ils étudiaient principalement: les processus automatiques d'une part et les processus contrôlés d'autre part. Nous avons présenté quelques comparaisons ou recoupements entre les résultats observés dans les études en potentiels évoqués et celles en TEP ou en IRMf sont présentées dans le chapitre 1.1.4.
1.1.3.1. Les potentiels évoqués et les processus attentionnels automatiques
La composante MMN et la détection automatique d'un changement dans l'environnement
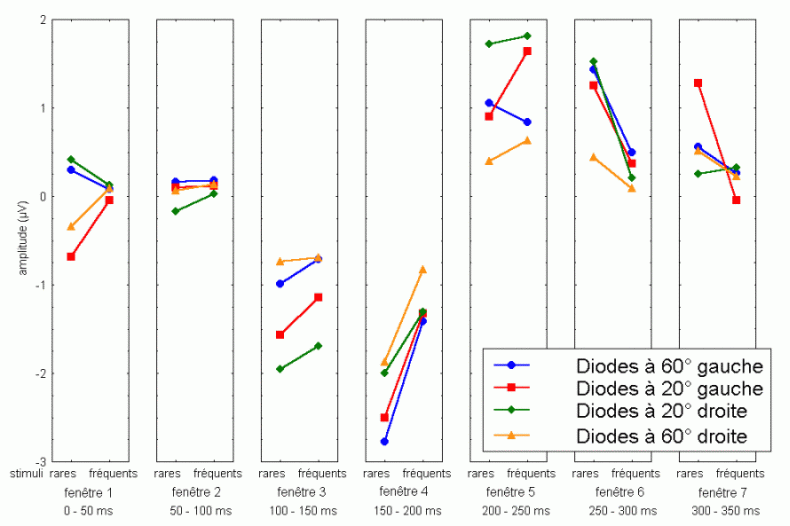
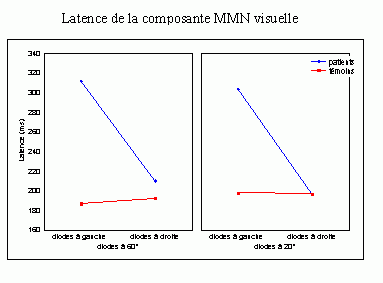
Les travaux de Näätänen et de ses collaborateurs sur les mécanismes attentionnels auditifs ont mis en évidence, notamment, une augmentation de la négativité du potentiel évoqué lorsque des stimuli fréquents (qualifiés de standards) et des stimuli rares (qualifiés de déviants) sont présentés successivement, dans un ordre aléatoire sans qu'aucune tâche ne soit demandée au sujet (Näätänen et al., 1978, pour une revue voir Näätänen, 1992). Dans une telle situation, on observe une augmentation de la négativité du potentiel évoqué par les stimuli rares par rapport au potentiel évoqué par les stimuli fréquents, environ 100 à 200 ms après la survenue des stimuli. Cette augmentation de la négativité correspond à la composante dénommée 'Mismatch negativity' ou 'MMN'. Cette composante a été largement étudiée dans la modalité auditive, sa présence dans d'autres modalités est encore incertaine. La première étude présentée dans notre propre travail (chapitre 6) a porté sur la recherche d'une telle composante dans la modalité visuelle.
Cette composante est sensible, notamment, aux différences (entre les sons rares et fréquents) de fréquence sonore (Sams et al., 1985 par exemple), aux différences d'intensité (Näätänen et al., 1978; Näätänen et al., 1987; Paavilainen et al., 1991; Woldorff et al., 1991), aux différences d'origine spatiale (Paavilainen et al., 1989), aux différences de durée (Paavilainen et al., 1993) et aux différences de changements phonétiques (Aaltonen et al., 1987). Cette composante est indépendante de la conscience qu'a le sujet de percevoir les stimuli déviants: elle apparaît même lorsque la différence entre les deux types de stimuli est trop faible pour être détectée consciemment ou lorsque le sujet est engagé dans une tâche principale (lire un livre par exemple) et qu'aucune réponse comportementale n'est exigée (Näätänen, 1992).
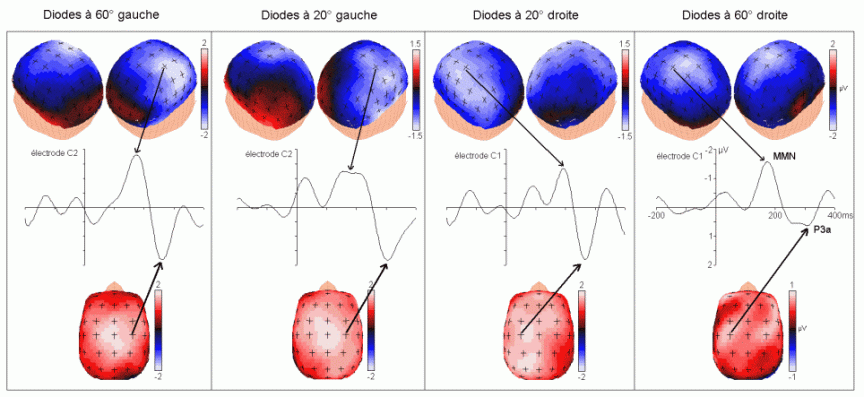
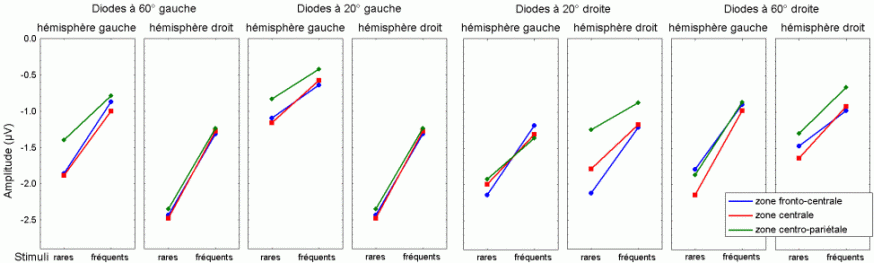
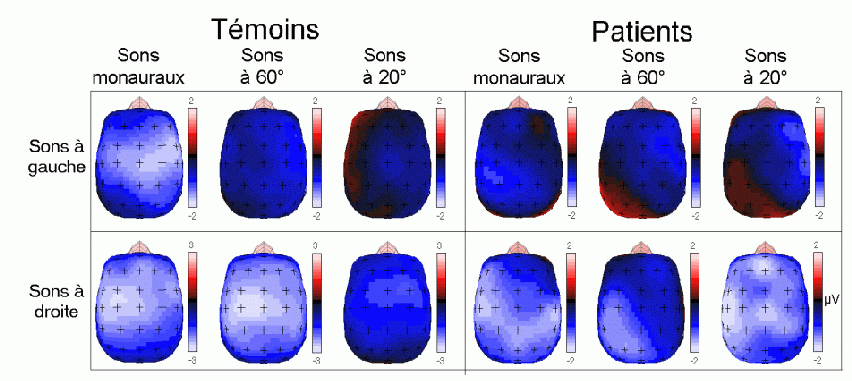
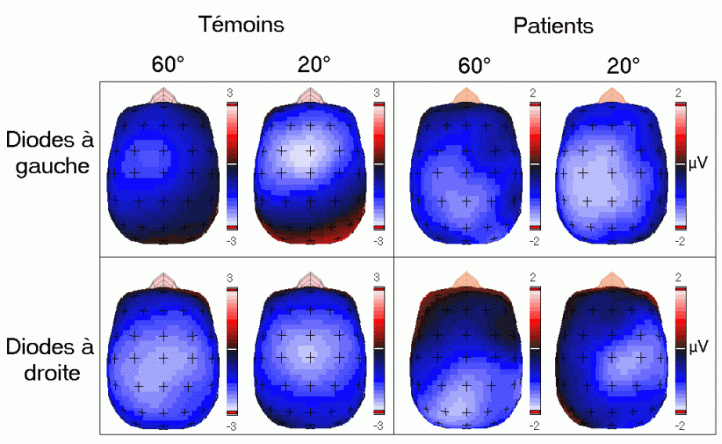
L'amplitude de la composante MMN culmine globalement dans les régions fronto-centrales (proche des électrodes FZ et CZ). La majorité des études sur cette composante ont mis en évidence une amplitude plus élevée sur l'hémisphère droit que sur l'hémisphère gauche, quelle que soit l'oreille stimulée. Les analyses de la distribution de la composante MMN sur le scalp et de ses générateurs sous-jacents suggèrent la présence d'une source dans chaque hémisphère, localisée au niveau du cortex auditif (Giard et al., 1990, Scherg et al., 1989). Les travaux de Giard et al. (1990), utilisant des cartographies de la densité de courant sur le scalp, ont suggéré que cette composante contienne effectivement des sources situées dans le cortex supra-temporal mais également dans les zones frontales. Dans une étude utilisant des électrodes implantées (Kropotov et al., 1995), les activités cérébrales sous-jacentes à la composante MMN ont été recueillies majoritairement au niveau des électrodes dans le cortex temporal, proche des aires 21 et 42 de Brodmann. Cependant, le générateur situé dans l'hémisphère controlatéral à l'oreille stimulée présenterait une amplitude plus élevée que son homologue situé dans l'hémisphère ipsilatéral (Giard et al., 1990). Ces résultats ont été confirmés par des études en magnétoencéphalographie (Hari et al., 1980) et par des enregistrements intra-cérébraux chez l'animal (Csépe et al., 1987, Javitt et al., 1995; Kropotov et al., 1995).
La source frontale (ou préfrontale selon les auteurs) serait présente uniquement dans l'hémisphère droit. La diminution de l'amplitude de la composante MMN chez de patients affectés de lésions situées dans le cortex préfrontal dorsolatéral (Alho et al., 1994) confirme l'implication du lobe frontal dans la génération de cette composante. Cette composante proviendrait donc d'une source sensorielle spécifique (temporale) dans chaque hémisphère et une source frontale droite. Ces sources (temporales et frontale) auraient des fonctions différentes selon Giard et al. (1990). Le processus de détection de l'apparition d'un stimulus différent, nouveau par rapport aux stimuli précédents, serait assuré au niveau des aires temporales. Selon Giard, ce processus déclencherait un mécanisme d'orientation automatique de l'attention assuré au niveau des aires frontales droites. La présence de cette composante témoignerait donc de l'existence d'un processus attentionnel automatique de détection d'un changement dans l'environnement.
La composante P3a et l'orientation automatique de l'attention
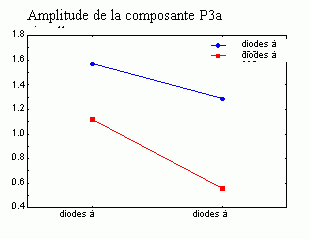
La composante MMN est souvent suivie par un pic positif situé dans les régions fronto-centrales, environ 230 à 350 ms après la présentation des stimuli rares (Näätänen, 1992). Cette positivité est généralement appelée composante 'P3a'. Elle se distingue selon plusieurs critères de la composante P3b, présentée dans le prochain chapitre. La composante P3a est consécutive à l'apparition d'un stimulus nouveau (inattendu, rare et incongru) alors que la composante P3b est observée lorsque le sujet porte son attention sur un stimulus cible. Les deux composantes se différencient également au niveau de leur latence et de leur topographie (respectivement plus précoce et plus frontale pour la composante P3a). Cette dernière a été mise en évidence dans les modalités auditive, visuelle et somesthésique (Courchesne et al., 1975; 1984; Knight, 1984; Renault et al., 1982; Yamaguchi et Knight, 1991 notamment).
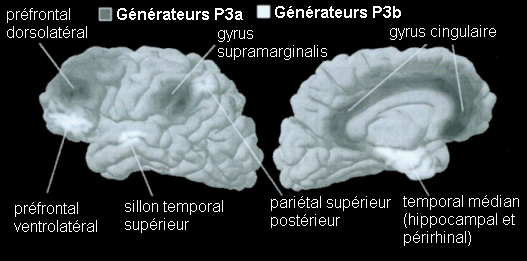
Les études portant sur les aires cérébrales sous-jacentes à cette composante ont mis en évidence des résultats légèrement différents (Knight, 1996; 1997; Polich et Squires, 1993; Halgren et al., 1995a,b). Dans une étude portant sur la modalité auditive, Knight et al. (1989) ont observé une diminution importante de l'amplitude de la composante P3a (et de la composante P3b) lors de lésions situées dans le cortex temporal. Selon ces auteurs, des lésions situées dans le lobe pariétal auraient également un effet sur cette composante. A partir des potentiels intra-cérébraux, Kropotov et al. (1995) ont observé au niveau de l'aire 42 de Brodmann (cortex temporal) une positivité succédant à la composante MMN. Cette positivité, observée également dans les cortex frontaux et pariétaux, pourrait être associée à la composante P3a. Lors d'une étude utilisant conjointement les techniques des potentiels évoqués et de l'IRMf, Opitz et al. (1999a, b) ont montré la présence de sources temporales sous-jacentes à cette composante. Une activité frontale lors d'une modélisation des générateurs cérébraux sous-jacents à cette composante a également été proposée par Mecklinger et Ullsperger (1995). A partir d'enregistrements intracérébraux, Halgren et al. (1998) suggèrent que les générateurs la composante P3a se situent dans le cortex pariétal inférieur, le cortex cingulaire et le cortex préfrontal dorsolatéral (Fig. 5).
Ces résultats semblent donc mettre en évidence l'implication de différentes zones cérébrales dans la génération de cette composante: des régions frontales et pariétales ainsi que des zones temporales. En revanche, la majorité des auteurs s'accordent sur le processus fonctionnel que refléterait cette composante: un changement d'orientation involontaire de l'attention vers un stimulus apparaissant d'une manière inattendue (Ford et al., 1976; Friedman et Simpson, 1994; Näätänen, 1992 notamment).
En résumé
Les études électrophysiologiques permettent donc d'étudier le décours temporel des processus attentionnels automatiques. Dès 120 ms environ après la présentation d'un stimulus nouveau et inattendu on observe un effet sur les potentiels évoqués. Ces processus seraient donc reflétés par les composantes MMN et P3a qui témoigneraient respectivement d'un processus involontaire de détection d'un changement dans l'environnement et d'un processus d'orientation automatique de l'attention. Ces composantes sont observées dans des situations de perception passive de stimuli rares et fréquents mais également dans les situations d'attention sélective. Cependant, dans la plupart de ces dernières situations, elles sont légèrement masquées par les composantes N2b et P3b, abordées dans le prochain chapitre. Les générateurs sous-jacents aux composantes MMN et P3a ne paraissent pas être encore clairement spécifiés. Il semble toutefois que les régions temporales droites et gauches et la région frontale droite participent à la génération de la composante MMN. Pour la composante P3a, les générateurs pourraient se situer dans les régions temporales, frontales et/ou pariétales.
1.1.3.2. Les potentiels évoqués et les processus attentionnels contrôlés
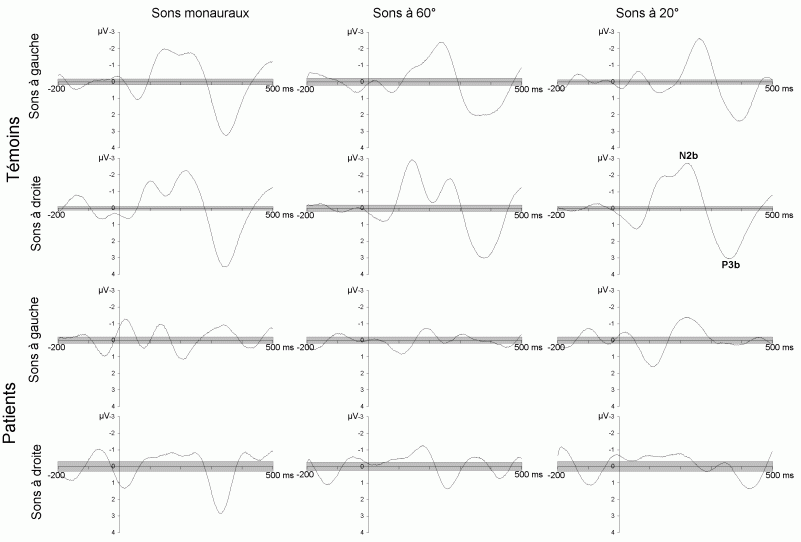
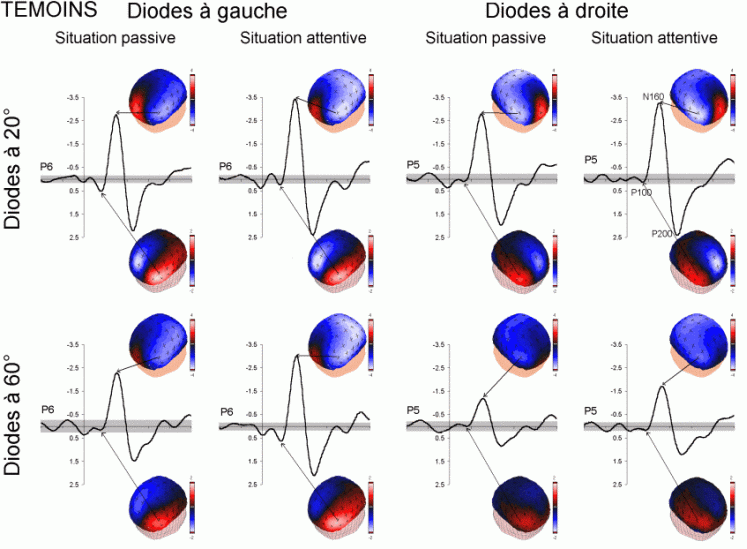
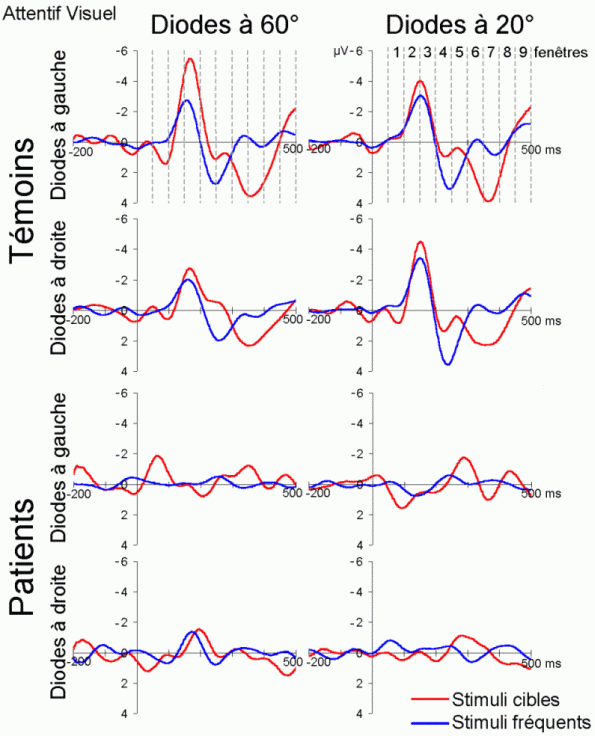
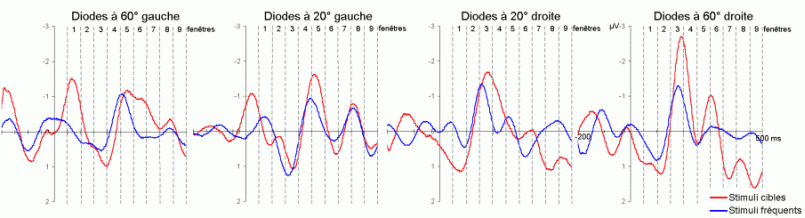
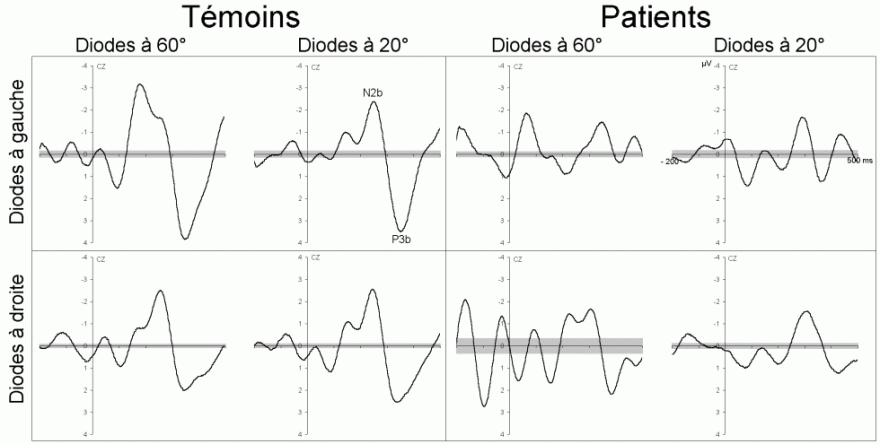
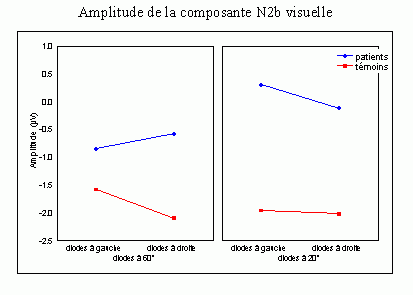
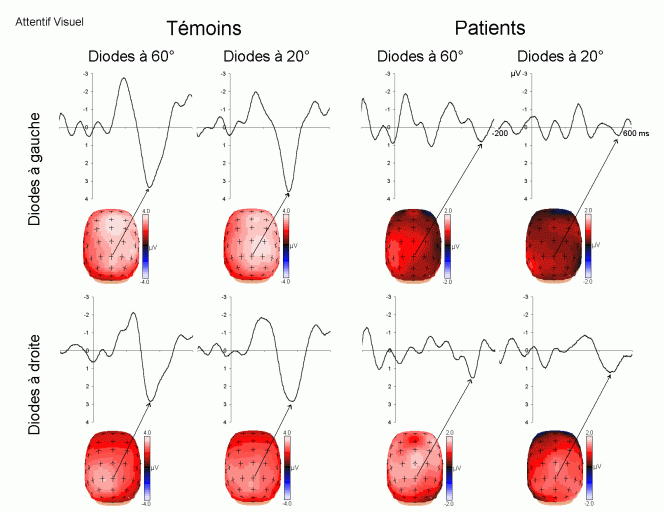
L'étude du décours temporel des mécanismes attentionnels contrôlés a également été effectuée à partir du paradigme 'oddball'. Ce paradigme permet effectivement d'observer certaines composantes (autres que les composantes MMN et P3a) lorsque le sujet doit porter son attention sur les stimuli rares (appelés 'stimuli cibles' dans ce cas). La tâche du sujet peut être par exemple de compter silencieusement les cibles qu'il perçoit ou d'appuyer sur un bouton-réponse. La différence entre les potentiels évoqués par les stimuli cibles et ceux évoqués par les stimuli fréquents permet d'isoler ce qui est lié spécifiquement à l'orientation volontaire de l'attention et à la détection de la cible. L'analyse de cette différence révèle une négativité fronto-centrale, appelée composante N2b, culminant entre 150 et 300 ms après la présentation des stimuli. Une positivité plus tardive apparaît ensuite, entre 300 et 600 ms, elle culmine au niveau des régions pariétales et est généralement appelée 'composante P3b'.
Ces composantes sont observées lorsque le sujet est engagé dans une tâche de discrimination de stimuli ou d'attention sélective, dans les modalités auditive et visuelle (Squires et al., 1975; Ritter et al., 1979; Renault et Lesèvre, 1979). D'autres composantes attentionnelles (voir Näätänen, 1992 pour une revue exhaustive) comme la 'négativité de différence' (Nd) ou la 'Processing Negativity' (PN) sont également observées dans des situations d'attention sélective (dans des paradigmes cependant légèrement différents). Nous présentons ci-dessous les travaux concernant les composantes N2b et P3b correspondant aux ondes que nous avons étudiées dans les résultats (chapitres 8 et 9).
Dans les tâches mettant en jeu des processus attentionnels, on observe un effet précoce sur les potentiels évoqués par les stimuli cibles avant l'apparition des composantes N2b et P3b. En effet, dans les modalités auditive et visuelle, environ 80 ms après la présentation des cibles cible, on peut observer une augmentation du potentiel sur les courbes évoquées par les stimuli cibles par rapport à celles évoquées par les stimuli fréquents (Hillyard et al., 1973; Harter et Aine, 1984; Hillyard et Muente, 1984). Cette augmentation a été souvent appelée 'l'effet N1'. Selon Hillyard et al. (1998), cette augmentation témoignerait d'un processus de gain sensoriel, ou d'amplification, au niveau des aires sensorielles traitant le stimulus attendu. Un tel processus sensoriel de gain a été également observé dans des études en TEP (Corbetta et al., 1991; Heinze et al., 1994; O'Leary et al., 1996). Dans une étude utilisant les potentiels évoqués et la TEP, Woldorff et al. (1997) ont suggéré que ce processus de gain se situe au niveau des aires visuelles V2, V3 ou V4 (Desimone, 1998). La localisation de ce processus au niveau de ces aires témoignerait de sa nature plus sensorielle qu'attentionnelle.
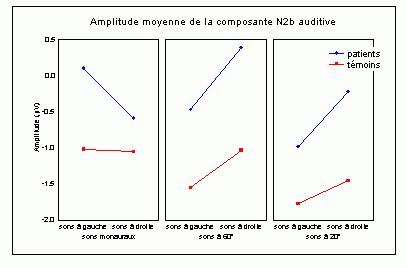
La composante N2b et l'orientation volontaire de l'attention
Des stimuli rares auxquels le sujet doit prêter attention entraînent des effets au niveau de l'amplitude des potentiels évoqués environ 200 ms après la présentation des ces stimuli. Une augmentation de la négativité, correspondant à la composante N2b est observée sur les potentiels évoqués par les stimuli cibles par rapport aux potentiels évoqués par les stimuli fréquents. Contrairement à la composante MMN, l'onde N2b est principalement présente lorsque le sujet porte son attention sur une cible donnée (Näätänen et Gaillard, 1983). Cependant, dans des situations passives (aucune tâche à effectuer), lorsque la différence entre ces stimuli rares et fréquents est très grande, cette composante peut également apparaître avec une faible amplitude. La topographie de cette composante n'est pas spécifique à une modalité: l'amplitude maximale se situe dans la région du vertex dans la modalité auditive comme dans la modalité visuelle (Näätänen, 1992). En revanche, les générateurs de cette composante ne sont pas encore actuellement bien spécifiés. Halgren et al. (1998) ont proposé que ces générateurs se situent dans le cortex rhinal. Kropotov et al. (1995) ont observé, à l'aide d'électrodes implantées, des activités cérébrales principalement au niveau du gyrus cingulaire antérieur, à des latences correspondant à l'apparition des composantes reflétant des processus attentionnels contrôlés (composantes N2 et P3). Beaucoup d'auteurs ont suggéré en effet que les générateurs de la composante N2b correspondraient en partie à ceux de la composante P3b (Fig. 5).
Différentes interprétations fonctionnelles de cette composante peuvent être observées dans la littérature. Selon Ritter et al. (1979), cette composante refléterait un processus de catégorisation du stimulus, alors que selon Näätänen et Gaillard (1983), cette onde témoignerait d'un processus d'éveil et d'orientation de l'attention vers la cible (Loveless, 1983, 1986). En accord avec cette dernière hypothèse, la plupart des auteurs semblent actuellement considérer que cette composante refléterait une attention orientée et contrôlée sur un stimulus donné (Renault et al., 1989; Näätänen, 1992).
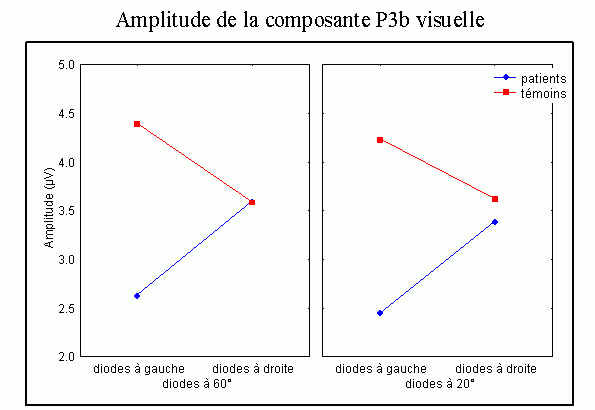
La composante P3b et l'identification des stimuli
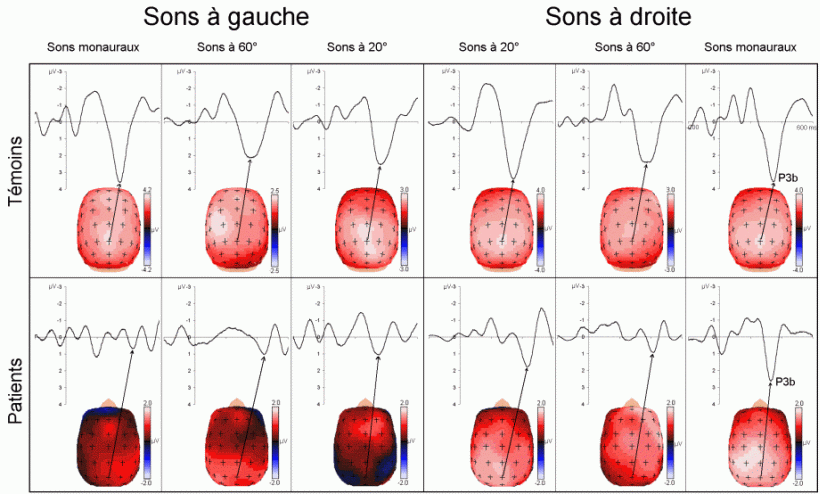
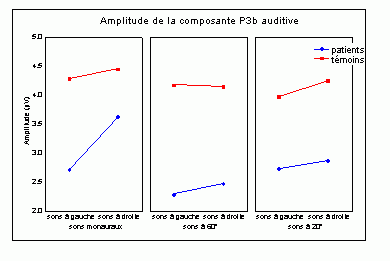
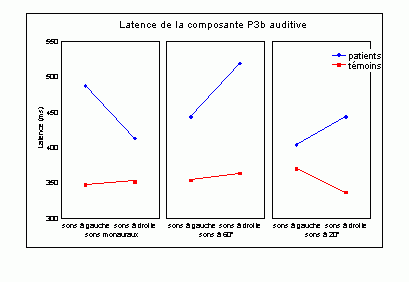
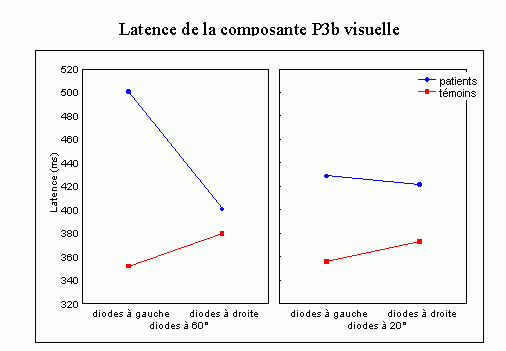
Sutton et al. (1965) ont observé une positivité assez tardive sur les potentiels évoqués visuels et auditifs qui a été appelée composante 'P300' (Vaughan et Ritter, 1970), P3 ou P3b (pour la différencier de la composante P3a). Cette composante est certainement l'onde qui a été la plus étudiée dans les modalités auditive, visuelle et somesthésique. Elle est constituée d'une positivité culminant dans la région pariéto-centrale (Smith et al., 1970, pour une revue voir par exemple Ducan-Johnson et Donchin, 1982; Picton, 1992) et peut souvent atteindre une grande amplitude (6-8 µV). Les caractéristiques de cette onde semblent actuellement bien connues. Elle est observée uniquement lorsque le sujet porte son attention sur le stimulus (Ritter et al., 1968). L'amplitude et la latence de cette composante varient en fonction de plusieurs facteurs tels que la probabilité d'apparition du stimulus ou la difficulté de la tâche. Plus la probabilité est faible, plus l'amplitude est élevée (Duncan-Johnson et Donchin, 1977). L'augmentation de la difficulté à discriminer les stimuli cibles des autres stimuli diminue l'amplitude et augmente la latence de cette composante (Squires et al., 1973).
Les enregistrements intracérébraux ainsi que les études de patients présentant des lésions cérébrales ont suggéré la présence de sources multiples pour la composante P3b (Fig. 5). Dans une étude utilisant conjointement les potentiels évoqués et l'IRMf, Linden et al. (1999) ont observé, à une latence correspondant à celle de la composante P3b, des activités dans le gyrus supramarginalis, dans le cortex insulaire ainsi que dans certaines régions plus restreintes (frontales et pariétales). Lors d'enregistrements intracérébraux, des activités similaires à cette composante ont été observées au niveau de l'amygdale, de l'hippocampe, du sillon temporal supérieur, du cortex orbitofrontal et du sillon intrapariétal (Halgren et al., 1980; 1998; Puce et al., 1989), dans les lobes frontaux et temporaux (Alain et al., 1989; Smith et al., 1990) ainsi que dans les ganglions de la base et les noyaux thalamiques (Kropotov et Ponomarev, 1991; Velasco et al., 1986). Ces résultats ont donc été observés dans les deux modalités.
Les travaux étudiant les effets de lésions cérébrales sur cette composante ont également mis en évidence l'implication de différentes régions dans la génération de la composante P3b (pour une revue voir Knight, 1990). Une diminution de l'amplitude de cette onde dans la modalité auditive a été mise en évidence lorsque les lésions se situent au niveau de la jonction temporo-pariétale et de la région frontale (Knight et al., 1989). Ces auteurs ont toutefois observé une composante d'amplitude 'normale' chez des patients affectés de lésions plus supérieures du lobe pariétal. Smith et Halgren (1988) ont également mis en évidence une diminution similaire de cette composante, dans la modalité visuelle, chez des patients affectés de lésions inféro-temporal (lobectomies).
Cependant, la contribution de chacune de ces aires aux processus d'identification des cibles n'est pas encore bien déterminée. Les études en imagerie fonctionnelle présentées ci-dessus ont suggéré que le cortex cingulaire antérieur soit impliqué dans des processus de détection et de sélection (Corbetta et al., 1991; Pardo et al., 1990; Posner, 1994). En plus du cortex cingulaire, le gyrus supramarginalis et le thalamus seraient aussi impliqués dans les processus de détection des cibles selon Menon et al. (1997). L'influence de la probabilité des stimuli suggère l'implication d'un processus d'attente ou de surprise. Selon certains auteurs, cette composante refléterait une mise à jour de la mémoire de travail (Donchin, 1981), pour d'autres auteurs elle témoignerait d'un processus d'identification de la cible (Neville et al., 1986). Il semble toutefois que les auteurs s'accordent, actuellement, sur l'hypothèse suggérant que la composante P3b reflète principalement des processus attentionnels contrôlés liés à l'identification de la cible.
En résumé
L'étude électrophysiologique des processus attentionnels contrôlés a mis en évidence deux composantes successives: les composantes N2b et P3b. Nous rappelons que d'autres composantes reflétant de tels processus ont également été étudiés dans la littérature (ondes Nd et PN). Les travaux que nous avons abordés ci-dessus mettent en évidence plusieurs résultats:
- Les composantes N2b et P3b ne seraient pas 'modalité-spécifiques': elles présentent des topographies similaires dans les modalités auditive et visuelle,
- La composante N2b refléterait un processus d'orientation volontaire de l'attention vers le stimulus attendu, actuellement ses générateurs ne sont pas encore clairement déterminés.
- Les zones cérébrales sous-jacentes à la composante P3b sont également partiellement indéterminées, plusieurs études ont mis en évidence des activités dans les régions temporo-pariétales-frontales, mais également dans d'autres aires corticales et sous-corticales.
1.1.4. Attention auditive et attention visuelle
La plupart des études dans le domaine de l'attention n'ont porté que sur une seule modalité sensorielle, généralement la modalité visuelle, et dans une moindre mesure, la modalité auditive. La présence de processus attentionnels qui pourraient être communs ou différents entre ces deux modalités reste encore actuellement en question. L'étude des processus attentionnels dans les modalités auditive et visuelle nécessite de connaître les différences anatomiques et fonctionnelles qui existent entre ces deux systèmes sensoriels. En effet, les différences entre les processus attentionnels auditifs et visuels pourraient être liés à des différences anatomiques entre les deux systèmes sensoriels. Avant d'aborder les travaux portant sur les aspects communs à ces deux modalités, nous avons décrit brièvement l'anatomie de ces systèmes. Nos études portant sur des tâches de détection et de localisation de cibles sonores ou visuelles, nous avons également présenté ci-dessous les processus de localisation spatiale observés dans chaque modalité.
1.1.4.1. Les systèmes sensoriels auditifs et visuels et leurs mécanismes de localisation spatiale
Le système auditif
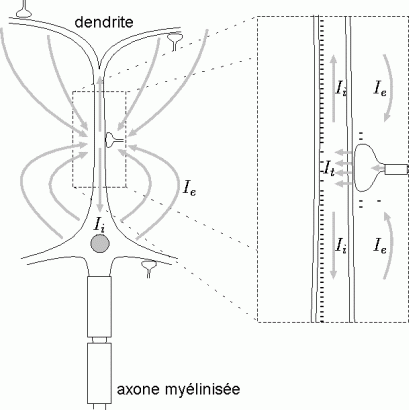
Une stimulation sonore provoque l'activation successive et/ou simultanée de différentes structures anatomiques du système sensoriel auditif (Fig. 6). La partie visible de ce système, l'oreille externe, est formée essentiellement de cartilage constituant le pavillon de l'oreille. Ce dernier permet de canaliser les sons à l'intérieur de la tête. Le passage vers l'oreille moyenne se fait par le canal auditif jusqu'au tympan. Une série de petits os, les osselets, est rattachée à la surface interne du tympan. Placés dans une cavité remplie d'air, les osselets transmettent les vibrations du tympan à une autre membrane, la fenêtre ovale. Derrière cette membrane, remplie de fluide (oreille interne), se trouve la cochlée qui génère une réponse nerveuse aux vibrations de la membrane de la fenêtre ovale. La cochlée contient en effet les cellules ciliées réceptrices. A ce niveau, on observe l'origine des fibres du nerf auditif. La réponse produite par le système auditif périphérique (oreilles externe, moyenne et interne) est transmise et analysée dans une série de noyaux du tronc cérébral (Fig. 6). Cette réponse parvient ensuite au thalamus, au niveau du corps genouillé médian. Depuis ces noyaux, les fibres se projettent sur le cortex auditif primaire situé dans le lobe temporal, sur la face supérieure de la première circonvolution temporale (gyrus transverse de Heschl, aires 41 et 42). Il faut également préciser que le système auditif contient d'autres voies de projections et de connexions secondaires entre différents noyaux qui n'ont pas été citées ci-dessus.
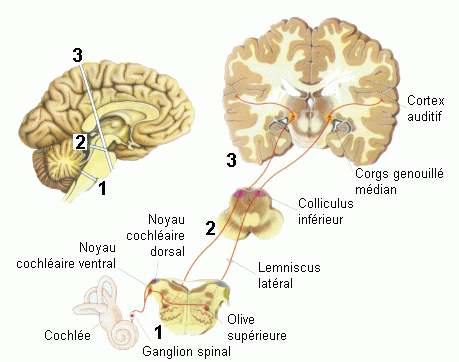
*Indication de plusieurs relais de ces voies. Les fibres auditives de chaque oreille se projettent dans le cortex gauche et dans le cortex droit.
Dans la cochlée, les cellules ciliées sont excitées par les déformations de la membrane basilaire. Chaque partie de la membrane est sensible (elle se déforme spécifiquement) à certaines fréquences. Tout le long des différents relais et dans le cortex auditif on retrouve une telle particularité qui est appelée 'tonotopie' (voir cependant Crottaz-Herbette et Ragot, 2000; Fig. 15 et 16). La tonotopie peut être définie comme une organisation spatiale des fréquences sonores. La discrimination des fréquences s'effectue donc à partir de la cochlée grâce à la sélectivité fréquentielle de la membrane basilaire, elle se maintient tout le long des relais à l'aide des filtres auditifs. Ces filtres correspondent à des mécanismes qui amplifient une certaine fréquence (valeur centrale du filtre) et atténuent les autres fréquences. Chaque neurone est particulièrement sensible à une fréquence donnée, appelée la fréquence caractéristique, et est moins sensible aux fréquences proches. Dans chaque relais, les neurones présentent également des propriétés différentes. Par exemple, les cellules dans l'olive supérieure sont activées par les noyaux cochléaires à partir des deux côtés du tronc cérébral. Ces neurones qui répondent à la stimulation des deux oreilles semblent jouer un rôle important dans la localisation des sons comme nous le présentons ci-dessous.
Une des propriétés importantes du système auditif est la projection sur chaque hémisphère des fibres auditives provenant de l'oreille droite et de l'oreille gauche (Fig. 6). La projection de ces fibres est toutefois plus importante sur l'hémisphère controlatéral à l'oreille stimulée (Rosenzweig, 1951). Une telle propriété n'est pas observée dans le système visuel qui possède une latéralisation complète de ses projections (projection controlatérale au champ visuel stimulé), comme nous le verrons dans la figure 7).
Les zones anatomiques responsables du traitement de toutes les caractéristiques des sons ne sont pas encore actuellement bien déterminées. L'analyse de la fréquence (tonotopie) et de la partie du spectre sonore impliquée semble s'effectuer (Crottaz-Herbette et Ragot, 2000) au niveau du cortex auditif primaire (aire 41 de Brodmann ou gyrus de Heschl), du cortex associatif primaire (aire 42) ou du cortex associatif secondaire (aire 22). Nous rappelons que nos études portent sur la détection de source sonore localisée à droite ou à gauche du sujet, nous présentons donc ci-dessous les principaux mécanismes de localisation sonore de notre système auditif.
La localisation sonore
Contrairement au système visuel qui possède une représentation spatiale du champ visuel (rétinotopie), le système auditif doit 'recréer' l'information spatiale à partir des caractéristiques des sons lui parvenant. Les mécanismes de localisation du son se basent sur plusieurs indices différents (Blauert, 1983). Le pavillon et les mouvements de tête jouent un rôle dans la localisation sonore. Les replis et les lobes de l'oreille externe réfléchissent et modifient le son. Ils permettent ainsi d'éviter, au moins partiellement, les confusions de localisation entre des sons provenant de l'avant ou de l'arrière. Les mouvements de tête quant à eux fournissent des informations supplémentaires en faisant varier les différences d'intensités et de délai entre les sons parvenant à chaque oreille. Il semble en effet qu'une bonne localisation repose principalement sur deux mécanismes complémentaires liés à la comparaison des sons atteignant les deux oreilles. L'importance relative de ces deux mécanismes varie en fonction de la fréquence considérée.
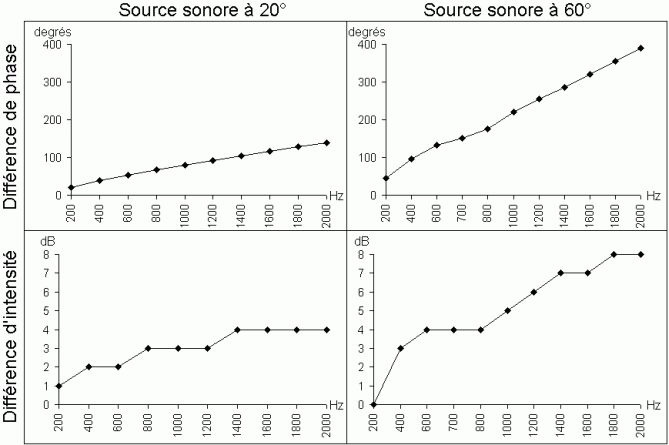
Le premier mécanisme repose sur la différence de temps d'arrivée du son entre les deux oreilles, différences appelées 'délai interaural'. Lorsque la source sonore ne se situe pas dans le plan médian, le son met plus de temps à parvenir à l'une des oreilles qu'à l'autre. Lorsque le début du son n'est pas perçu, ou lorsque le son est continu, la seule comparaison possible entre les deux oreilles porte sur la phase de l'onde sonore parvenant à chaque oreille. Cependant, lorsque les fréquences sont hautes (plus de 2000 Hz notamment), le délai interaural ne permet plus de localiser le son: la longueur d'onde est plus petite que la distance entre les deux oreilles. Cela signifie que plusieurs cycles peuvent se succéder entre le moment où le signal atteint une oreille puis l'autre. Ainsi, la comparaison des phases des signaux n'apporte plus d'information sur le délai temporel entre les signaux, et donc, sur la localisation de la source sonore.
Lorsque les sons ont des fréquences élevées, le cerveau se base sur un autre mécanisme dans la localisation sonore: la différence d'intensité interaurale. La tête représente une sorte d'écran pour la propagation du son et entraîne une différence d'intensité entre les signaux parvenant à chaque oreille. Il existe une relation directe entre la direction d'origine du son et l'effet de la tête sur l'intensité des signaux arrivant à chaque oreille. Pour un son situé sur la droite (à 90°), l'intensité sera nettement plus faible au niveau de l'oreille gauche qu'au niveau de l'oreille droite, et vice-versa lorsque le son est situé sur la gauche. En revanche, si le son vient d'en face (0°), les intensités des signaux parvenant à chaque oreille seront similaires; si la source sonore se situe légèrement sur la droite (par exemple 20°), le signal parvenant à l'oreille gauche sera légèrement moins intense que le signal parvenant à l'oreille droite. Ces différences d'intensité permettent aux neurones sensibles à l'intensité de localiser le son. Cependant, l'analyse de ces différences ne met pas en évidence d'effets nets lorsque les sons ont une fréquence faible (inférieure à 2000 Hz). Pour ces fréquences, les ondes sonores sont en effet diffractées tout autour de la tête et l'intensité des sons parvenant à chaque oreille est presque la même.
Il existe donc plusieurs processus de localisation des sources sonores qui sont donc différents et complémentaires. Pour des sons ayant une fréquence de 20 à 2000 Hz, le processus de localisation se baserait essentiellement sur le délai interaural. Lorsque les sons ont des fréquences plus élevées (de 2000 à 20000 Hz), la différence interaurale d'intensité serait le mécanisme principal. Ces deux processus forment ce qui est généralement appelé la 'théorie de la double localisation' (Lindsay et Norman, 1977; Canévet, 1989). Comme nous l'avons présenté ci-dessus, certains neurones auditifs à partir de l'olive supérieure présentent la particularité d'être binauraux: ils répondent aux sons provenant des deux oreilles. Il est probable que les mécanismes de comparaison de la phase et de l'intensité que nous venons de décrire reposent sur les neurones binauraux présents dans l'olive supérieure. Lors de la construction des stimuli sonores, nous nous sommes basés sur ces deux mécanismes principaux de localisation du son.
Le système visuel
La perception du monde commence au niveau de la rétine. Cette dernière se trouve sur la partie arrière de l'oeil, elle est composée de photorécepteurs spécialisés dans la conversion de l'énergie lumineuse en activité nerveuse. La rétine montre une propriété importante: la rétinotopie. Cette propriété correspond à la conservation au niveau de la rétine de l'organisation spatiale du champ visuel. Elle est également observée lors des différents relais sensoriels et au niveau du cortex visuel primaire.
Cependant, le champ visuel n'est pas simplement projeté sur chaque rétine (Bear et al., 1999). Le champ visuel complet est l'espace (calculé en degrés) visible quand les yeux regardent droit devant (Fig. 7). L'espace gauche (ou droit), appelé également hémichamp ou champ visuel gauche (ou droit) correspondant à tout l'espace situé à gauche (ou à droite) d'un plan vertical passant par un point situé juste en face du sujet. L'hémichamp visuel gauche est reproduit à la fois sur la rétine nasale de l'oeil gauche (moitié droite de la rétine gauche) et sur la rétine temporale de l'oeil droit (moitié droite de la rétine droite), et vice-versa pour l'hémichamp visuel droit. La partie située au centre du champ visuel est perçue par la rétine gauche et par la rétine droite, cet espace est souvent appelé le champ visuel binoculaire. Les axones des neurones visuels se rassemblent pour former les nerfs optiques partent de la rétine en direction du chiasma optique (Fig. 7). A l'endroit d'où partent ces fibres il n'y a pas de cellules réceptrices, cette correspond à ce qui est appelé la tâche aveugle. Les fibres nerveuses de la partie nasale de la rétine gauche et celles de la partie temporale de la rétine droite passent du côté droit au niveau du chiasma optique, et vice-versa pour la partie nasale de la rétine droite et la partie temporale de la rétine gauche. Le croisement des nerfs optiques au niveau du chiasma optique a pour conséquence que l'hémichamp visuel gauche soit traité dans l'hémisphère droit, et l'hémichamp visuel droit soit traité dans l'hémisphère gauche.
Chez l'homme, le nerf optique, après le chiasma optique, se divise en deux grandes voies. Une de ces voies (90% des fibres optiques) conduit au corps genouillé latéral (partie dorsale du thalamus) par le tractus optique dont les axones se prolongent jusqu'au cortex visuel primaire correspondant à l'aire 17 de Brodmann, appelée également cortex strié ou aire V1 (Fig. 7). Ce dernier est situé dans le lobe occipital, c'est à ce niveau que débutent les analyses principales des évènements perçus visuellement: analyses des couleurs, de la forme, du mouvement et de la profondeur. La seconde voie (non illustrée sur la figure 7) conduit au colliculus supérieur (zone située au-dessus du tronc cérébral). La voie de la perception visuelle est celle qui passe par le corps genouillé latéral et qui aboutit au cortex visuel. Nous précisons à nouveau que les cellules du cortex strié présentent également une organisation rétinotopique.
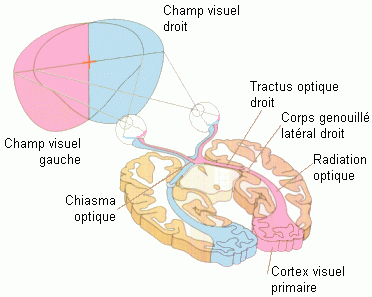
Les différents processus de traitement de l'information visuelle (traitement de la couleur, de la forme, du mouvement,...) commencent dès la rétine mais sont principalement élaborés dans le cortex strié et au-delà, dans d'autres zones du cerveau reliées les unes aux autres par de nombreuses connexions cortico-corticales. Ces processus seraient constitués de deux grands systèmes corticaux partant du cortex strié: un système s'étendrait vers le lobe pariétal et l'autre système se projetterait sur le lobe temporal. Le système pariétal contiendrait notamment l'aire V5 (ou MT) qui serait spécialisée principalement dans le traitement du mouvement. Le système temporal contiendrait l'aire V4, qui semble jouer un rôle fondamental dans la perception de la forme et de la couleur (Bear et al., 1999).
Différences entre les systèmes sensoriels auditif et visuel
Nous rappelons que nos études ont eu pour but, notamment, de comparer les processus attentionnels dans la modalité auditive et dans la modalité visuelle. La présentation des caractéristiques anatomiques de chaque système était destinée à mettre en évidence leurs aspects communs ou différents.
L'organisation fonctionnelle du système visuel semble donc actuellement mieux connue que celle du système auditif. En effet, les zones cérébrales sous-jacentes au traitement visuel de la couleur, de la forme et du mouvement semblent plus ou moins bien déterminées. L'organisation fonctionnelle connue pour le moment dans la modalité auditive concerne principalement le traitement des fréquences sonores avec la tonotopie. Des études utilisant la technique des potentiels évoqués ont cependant également mis en évidence la présence d'une organisation spatiale pour le spectre sonore (Crottaz-Herbette et Ragot, 2000, voir chapitre 3.2.3.2.).
D'autres différences existent entre les deux systèmes. Les processus de localisation spatiale semblent être plus complexes pour le système sensoriel auditif que pour le système sensoriel visuel. Le système visuel possède une représentation spatiale de champ visuel, la rétinotopie, présente au niveau de la rétine, dans certains noyaux sous-corticaux et dans le cortex visuel primaire. En revanche, la cochlée ne transmet pas d'information spatiale mais des informations fréquentielles (tonotopiques) et temporelles qui permettent de reconstruire une information spatiale. Finalement, une des différences évidentes entre les deux systèmes porte sur l'étendue du champ perceptif qu'ils peuvent prendre en compte. En effet, le champ visuel que nous pouvons percevoir spontanément, sans mouvement de la tête, se situe devant et sur les côtés. Il a un angle total d'environ 180°. En revanche, dans la modalité auditive, nous pouvons percevoir des sons venant de n'importe où, le champ perceptif auditif a donc un angle de 360° (le champ auditif est également plus étendu verticalement que le champ perceptif visuel).
En résumé
Ces constatations mettent en évidence des différences entre les deux modalités au niveau de leurs organisations anatomique et fonctionnelle. Les différences au niveau des mécanismes de localisation spatiale entre la vision et l'audition sont importantes à préciser dans ce travail puisqu'elles conduisent à des fonctionnements différents (le système auditif doit reconstruire une information spatiale alors que le système visuel possède une organisation spatiale directement). Finalement, il faut également préciser que l'organisation des différentes aires sous-jacentes au traitement de l'information visuelle est mieux déterminée que celle concernant l'information auditive.
1.1.4.2. Les processus attentionnels auditifs et visuels
Dans la vie quotidienne on peut facilement constater la présence de relations entre la vision et l'audition. Dans une salle de cinéma, nous avons l'impression que les paroles entendues sont prononcées par les acteurs sur l'écran alors que les sources sonores ne correspondent pas à l'emplacement des acteurs. La ventriloquie est un exemple de ces liens entre ces deux modalités: bien que les sources sonores et visuelles ne se situent pas au même endroit, nous 'fusionnons' les informations de ces deux sources en considérant qu'elles proviennent de l'endroit où se situe la source visuelle (Witkin et al., 1952). Un autre exemple de liens entre la vision et l'audition est notre tendance à tourner notre regard, d'une manière souvent involontaire et automatique, en direction d'un bruit soudain. Il semble donc qu'il existe des liens entre les processus attentionnels appliqués à des modalités différentes. On peut alors se demander si les processus attentionnels sont similaires en audition et en vision et si les zones cérébrales sous-jacentes à ces processus sont communes ou différentes entre les modalités. Ce chapitre permet d'aborder ces questions centrales dans nos travaux et dans la littérature actuelle portant sur l'attention. Nos travaux portent donc sur la présence de liens d'influence entre les processus attentionnels dans différentes modalités d'une part et sur l'existence de bases neurales communes ou propres à ces processus d'autre part. Nous avons choisi d'étudier plus spécifiquement l'attention spatiale ainsi que les processus attentionnels automatiques et contrôlés.
Ce chapitre se divise en deux parties. La première partie présente des résultats en électrophysiologie et en imagerie fonctionnelle ayant suggéré, selon les études, des différences ou des similitudes entre les processus attentionnels en vision et en audition. Cette partie porte également sur la présence éventuelle de zones cérébrales qui seraient communes aux processus attentionnels auditifs et visuels (zones 'multimodales'). La seconde partie porte sur la présence d'effets attentionnels intermodaux dans ces deux modalités. Un bref résumé de ces observations est présenté à la fin de ce chapitre.
Effets attentionnels similaires ou différents dans les deux modalités
L'analyse des similitudes et des différences entre les processus attentionnels auditifs et visuels a été abordée dans des études comportementales, dans des études utilisant les potentiels évoqués ainsi que dans des travaux en TEP et en IRMf. Dans des tâches d'attention sélective visuelle et auditive, Woods et al. (1992), Alho et al. (1992) et Eimer et Schröger (1998) ont observé des différences entre les deux modalités au niveau de la distribution sur le scalp des potentiels évoqués attentionnels auditifs et visuels (reflétant des processus contrôlés). Teder-Sälejärvi et al. (1999) ont confirmé la présence de tels effets tout en montrant également l'existence d'effets intermodaux (présentés dans le chapitre suivant). Les composantes Nd et PN ont donc été observées dans les deux modalités. Selon ces études, les différences entre les deux modalités, reflétées principalement sur les composantes Nd et PN, suggèreraient que les zones cérébrales sous-jacentes à ces composantes ne seraient pas similaires pour ces deux modalités (zones 'modalité-spécifiques').
De plus, certains travaux en TEP et en IRMf ont suggéré que des effets précoces de l'attention sur les processus de traitement perceptif soient similaires dans les modalités auditive et visuelle (mais situés dans des zones différentes). En effet, dans une étude utilisant la TEP lors d'une tâche d'attention sélective auditive, Tzourio et al. (1997) ont observé une augmentation d'activité au niveau des lobes temporaux gauche et droit plus importante dans l'hémisphère controlatéral à l'oreille sur laquelle l'attention se porte (Fig. 8). O'Leary et al. (1986) ont observé un résultat similaire en IRMf. Dans une étude portant sur l'attention spatiale visuelle, et utilisant conjointement les techniques des potentiels évoqués et de la TEP, Heinze et al. (1994) ont montré une augmentation d'activité dans l'hémisphère controlatéral au côté de présentation des cibles. Plus précisément, lorsque l'attention portait sur le côté gauche, ces auteurs ont observé une activité au niveau du cortex visuel extrastrié droit (gyrus fusiforme postérieur), et quand l'attention se concentrait sur la droite, une activité au niveau des même zones mais dans l'hémisphère gauche a été constatée. Dans une étude portant sur l'attention spatiale visuelle, Corbetta et al. (1991) ont observé des augmentations d'activités au niveau des aires cérébrales sous-jacentes au traitement de la forme ou de la couleur selon le type de caractéristiques de la cible sur lequel le sujet doit porter son attention (forme ou couleur). Les zones cérébrales responsables de l'analyse perceptive de certaines caractéristiques du stimulus visuel sont donc modulées par l'attention.
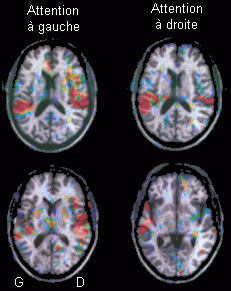
*En haut, activités bilatérales situées au niveau de la région du gyrus de Heschl. En bas, activités plus importantes dans l'hémisphère controlatérale à l'oreille attentive au niveau du gyrus temporal supérieur
Ces résultats montrent que l'attention entraîne des modifications d'activités localisées dans des aires responsables des traitements perceptifs de l'information. Tzourio et ses collaborateurs suggèrent que l'augmentation qu'ils observent dans la région postérieure du gyrus temporal supérieur est similaire à ce qui est observé en potentiel évoqué lors de 'l'effet N1' (Hillyard et al., 1973, voir le chapitre 1.1.3.2.). Cette suggestion peut également concerner les résultats de Heinze et al., (1994) au niveau du cortex visuel extrastrié puisque l'effet 'N1' a été également observé dans la modalité visuelle et dans la modalité somesthésique (Näätänen, 1990). Ces asymétries pourraient provenir de l'anatomie des voies sensorielles qui se projettent majoritairement (voies auditives) ou complètement (voies visuelles) dans l'hémisphère controlatéral au côté de présentation des stimuli. Dans la modalité auditive comme dans la modalité visuelle, l'attention pourrait influencer précocement le traitement perceptif des stimuli (dès 80 - 130 ms dans l'étude de Heinze) en 'augmentant' l'activité cérébrale des zones responsables de l'analyse des traits sur lesquels l'attention se porte spécifiquement (par exemple la localisation spatiale ou la couleur du stimulus). Ces études suggèrent donc l'existence de processus attentionnels précoces qui seraient similaires dans les deux modalités.
L'ensemble de ces études met en évidence des similarités entre les processus attentionnels en vision et en audition (les composantes Nd et PN ont été observées dans les deux modalités) alors que les zones cérébrales sous-jacentes à ces processus seraient différentes entre les deux modalités. Cependant, d'autres études en potentiels évoqués, en TEP et en IRMf parviennent à des résultats différents suggérant des bases neurales communes.
Selon Halgren et al. (1998), la composante P3a (Fig. 5), ainsi que l'ensemble des composantes évoquée par la présentation d'un stimulus nouveau et inattendu, ne seraient pas 'modalité-spécifiques'. Ces auteurs suggèrent également que les composantes N2b et P3b, reflétant des processus conscients et contrôlés, reflètent l'activité de générateurs également similaires en audition et en vision. Comme nous l'avons mentionné ci-dessus à propos de la composante P3b (chapitre 1.1.3.2., figure 5), des activités ont été effectivement observées en vision et en audition dans le gyrus supramarginalis, dans le cortex insulaire et dans des régions frontales et pariétales (Linden et al., 1999). D'autres régions cérébrales participeraient à la génération de cette composante (Smith et al., 1990; Halgren et al., 1995a, b; Baudena et al., 1995; Knight et al., 1989). La composante N2b présente des topographies similaires en vision et en audition (Halgren et al., 1998; Näätänen, 1992) qui suggèrent des bases neurales communes dans les deux modalités Ces composantes refléteraient des processus attentionnels automatiques ou contrôlés (selon la composante) qui semblent donc, selon ces études, avoir des bases neurales communes. Les générateurs sous-jacents à ces composantes se situeraient donc dans des zones multimodales, contenant au moins des informations auditives et visuelles. Cependant, la présence de zones communes à plusieurs modalités peut provenir soit de l'existence d'un processus multimodal, soit de la présence de multiple zones 'modalité-spécifiques' proches les unes des autres. Des études sur l'animal ont permis de déterminer la présence d'aires spécifiquement multimodales.
Des études sur le singe et le chat, principalement, ont suggéré en effet l'implication de zones multimodales dans l'attention spatiale notamment. Ces zones incluraient le cortex pariétal inférieur, des aires intrapariétales, le putamen, le cortex inférieur prémoteur ainsi que le colliculus supérieur (Andersen et al., 1997; Graziano et Gross, 1993; Stein et al., 1993). Les études de patients présentant des lésions cérébrales ont apporté également des informations sur de telles zones. Farah et al. (1989), notamment, ont mis en évidence, chez des patients affectés de lésions cérébrales localisées dans le lobe pariétal, des déficits dans la modalité auditive comme dans la modalité visuelle.
Les zones cérébrales sous-jacentes aux processus attentionnels semblent donc être nombreuses et leur caractère 'modalité-spécifique' ou 'multimodal' n'est pas encore clairement spécifié. D'après ces études, les processus attentionnels précoces seraient plutôt similaires dans les deux modalités mais ils proviendraient de zones différentes pour la vision et pour l'audition. De plus, les effets attentionnels plus tardifs observés sur les potentiels évoqués semblent provenir de plusieurs zones cérébrales dont certaines seraient propres à chaque modalité alors que d'autres seraient communes à différentes modalités. Les processus attentionnels observés en vision et en audition seraient donc globalement similaires tout en présentant des bases cérébrales communes ou différentes. De plus, ces processus ont des influences les uns sur les autres, comme le suggèrent les études portant sur l'intermodalité des processus attentionnels.
Effets attentionnels intermodalitaires
Dans une tâche d'indiçage proche de celle de Posner (présentée ci-dessus dans le chapitre 1.1.1.1.), Buchtel et Butter (1988) ont étudié l'influence de la présentation d'indices visuels ou auditifs (indices valides, non-valides ou neutres) sur la détection de cibles visuelles ou auditives latéralisées à droite ou à gauche. Ces auteurs ont montré que des indices valides auditifs ou visuels amélioraient les performances de détection des cibles visuelles. Bédart et al. (1993) ont confirmé ces résultats, et ont également montré une détérioration des performances lorsque l'indice n'indique pas l'emplacement correct de la cible (auditive ou visuelle). Des effets similaires entre les modalités tactile et visuelle ont été également mis en évidence (Butter et al., 1989). Ces études comportementales (pour une revue voir Driver et Spence, 1998) suggèrent la présence de liens d'influences réciproques entre les modalités (auditive, visuelle et tactile) comme le témoigneraient les effets d'indiçage spatial. L'orientation de l'attention dans une modalité peut donc être déclenchée et guidée par des indices provenant d'une autre modalité.
Les travaux de Alho et al. (1997), utilisant les potentiels évoqués, ont montré également l'existence de liens d'influence entre deux modalités, dans une tâche d'attention sélective visuelle durant laquelle les sujets entendaient des stimuli auditifs rares ou fréquents. La présentation des stimuli rares entraînait classiquement l'apparition d'une composante MMN sur les potentiels évoqués (suivie par une composante P3a) mais également une baisse des performances de détection des cibles visuelles. La présence de la composante MMN témoignerait du déclenchement de processus attentionnels automatiques suite à la survenue des stimuli auditifs rares. Ces processus entraîneraient une orientation de l'attention sur ces stimuli comme le refléterait la présence de la composante P3a. Un petit changement auditif peut donc entraîner une orientation involontaire de l'attention dans une modalité, même quand cette dernière est impliquée dans une tâche nécessitant une grande attention dans une autre modalité. Teder-Sälejärvi et al. (1999) ont étudié également les effets intermodaux à l'aide des potentiels évoqués. Ces auteurs ont observé une augmentation de l'amplitude des potentiels évoqués par des stimuli présentés dans la modalité inattendue se situant cependant à un emplacement attendu.
Ces travaux ont confirmé, à l'aide d'indices électrophysiologiques, les résultats d'études comportementales ayant montré que l'attention spatiale facilite le traitement des stimuli situés à l'endroit sur lequel l'attention se porte, même si le stimulus en question n'est pas dans la modalité attendue (par exemple Buchtel et Butter, 1988). Ces études montrent l'existence de liens entre les processus attentionnels dans différentes modalités. Ces liens pourraient témoigner de la présence de processus gérant les ressources attentionnels de plusieurs modalités. Il s'agirait de processus dont les zones cérébrales sous-jacentes seraient 'a-modales' ou multimodales. Plusieurs études, notamment chez l'animal, ont en effet émis l'hypothèse de l'existence de telles zones.
1.1.4.3. Conclusion
L'ensemble de ces études suggère certaines conclusions concernant les processus attentionnels auditifs et visuels mais laisse également en suspens plusieurs questions. Ces processus semblent fonctionner d'une manière assez similaire en audition et en vision, les composantes attentionnelles étant observées dans les deux modalités. Nous rappelons cependant que la composante MMN, reflétant un processus attentionnel automatique (chapitre 1.1.3.1.) n'a pas encore été clairement identifiée dans la modalité visuelle. La première étude que nous présentons ci-dessous porte précisément sur la recherche de cette composante en vision (chapitre 6).
La présence de zones cérébrales qui seraient responsables de traitements concernant plusieurs modalités est encore actuellement en question, notamment en ce qui concerne les processus attentionnels. Certaines études montrent des bases neurales communes aux deux modalités alors que d'autres suggèrent des aires cérébrales propres à chaque modalité. D'après l'ensemble de ces résultats, les zones cérébrales sous-jacentes aux processus attentionnels précoces seraient propres à chaque modalité (généralement localisées proches des aires 'perceptives') alors que les aires sous-jacentes aux processus plus tardifs seraient plus nombreuses, certaines seraient communes aux deux modalités et d'autres propres à chaque modalité. Cependant, les modèles de l'attention qui sont présentés ci-dessous ne distinguent pas les modalités auditive et visuelle. Ils présentent le fonctionnement général de l'attention en spécifiant les différents processus et les réseaux neuronaux sous-jacents sans différencier les modalités entre elles.
1.3. Les modèles de l'attention
La littérature sur l'attention contient beaucoup de modèles, nous n'en avons cependant abordé que quelques uns dans ce présent travail. Dans la première partie de ce chapitre, nous avons rappelé brièvement les concepts sous-jacents à certains des principaux modèles de l'attention (modèles de Broadbent, de Treisman, et de Schneider et Shiffrin). Nous avons ensuite présenté d'une façon plus détaillée d'autres modèles de l'attention qui ont tenté de rattacher les processus attentionnels qu'ils décrivent à des réseaux et des structures cérébrales (modèles de Laberge, de Posner et de Mesulam).
1.3.1. Les théories du filtrage: le modèle de Broadbent
Broadbent est certainement l'un des premiers auteurs à avoir étudié les effets de l'attention sélective (Broadbent, 1958; 1970). Il a considéré l'attention sélective essentiellement comme un processus de filtrage permettant d'éviter que le système global ne soit surchargé. Tous les stimuli seraient encodés et analysés en parallèle au niveau de leurs caractéristiques physiques (à un niveau de traitement préattentif). Cependant, pour que les informations soient réellement 'perçues', elles doivent être sélectionnées une à une. Ces processus de filtrage seraient basés sur les caractéristiques physiques générales de l'information. L'origine spatiale, le timbre ou l'intensité d'une voix seraient par exemple des aspect traités précocement par le système auditif. Ces mécanismes sensoriels seraient situés en amont des opérations cognitives elles-mêmes. La sélection simultanée de plusieurs informations entraînerait une division de l'attention qui conduirait à un certain coût: chaque élément serait moins bien traité que s'il était le seul à avoir été sélectionné. D'après ce processus de filtre, les stimuli attendus (parvenant à l'oreille attentive par exemple) feraient l'objet de traitements supplémentaires alors que les stimuli non attendus (parvenant à l'oreille non attentive par exemple) seraient bloqués. Selon Broadbent, l'attention sélective agirait donc comme un filtre permettant la sélection des informations pertinentes et l'élimination, ou le blocage des informations non pertinentes.
Le modèle de Broadbent fut rapidement remis en question. Des études utilisant des tâches d'écoute dichotique montrèrent que les informations parvenant à l'oreille inattentive faisaient l'objet d'un traitement plus poussé que ce que Broadbent (1958) proposait dans son modèle. Les sujets pouvaient par exemple reconnaître leur nom quand il était prononcé dans l'oreille inattentive (Moray, 1959). La présence d'un traitement cognitif des items rejetés était difficilement conciliable avec la théorie de Broadbent postulant une sélection périphérique.
1.3.2. Les théories de la sélection tardive: le modèle de Treisman
Treisman proposa plusieurs modifications de ce modèle en suggérant principalement que la fonction de 'filtre' de l'attention serait plutôt un effet modérateur (Treisman, 1969). Le processus de filtre n'entraînerait pas un blocage du traitement des stimuli inattendus ou rejetés mais une atténuation de leur traitement. Ces processus permettraient donc de rendre compte de la détérioration des performances pour l'oreille inattentive (qui ne ferait pas l'objet d'un traitement très poussé), et de l'amélioration des performances pour l'oreille attentive (qui bénéficierait de toutes les ressources de traitement). Dans ce modèle, le rôle de l'attention se situerait donc à l'étape finale de la perception. Selon la théorie de l'intégration attentionnelle des 'attributs' ou des 'traits' proposée par Treisman (Treisman et Gelade, 1980), l'attention serait un 'ciment' qui relierait entre eux les différents codages perceptifs des dimensions (les traits) d'un objet.
Ces premiers modèles de l'attention se basaient essentiellement sur les expériences d'écoute dichotique. Les études sur l'attention effectuées depuis les années 70 privilégièrent de plus en plus la modalité visuelle en développant notamment des paradigmes d'amorçage (sémantique par exemple) et d'orientation visuo-spatiale de l'attention (voir le modèle de Posner présenté ci-dessous). Les travaux de Deutsch (notamment Deutsch, 1977) par exemple sur la perception tachistoscopique des lettres et les phénomènes d'amorçage ont permis de mettre en évidence la présence de traitements plus ou moins automatiques, qui seraient 'pré-attentifs', involontaires et effectués hors de la conscience. Ces travaux, entre autres, ont suggéré la présence d'une sélection tardive opérée par l'attention en fonction de l'utilité du stimulus. Cette sélection interviendrait après une série de traitements perceptifs automatiques pouvant avoir lieu en parallèle les uns des autres. Les études de Schneider et Shiffrin ont également proposé l'existence de traitements automatiques, mais également de traitements contrôlés, comme nous le décrivons ci-dessous.
1.3.3. La théorie des processus attentionnels automatiques et contrôlés
Un des modèles ayant eu d'importantes répercussions au niveau des théories en psychologie est celui de Scheider et Shiffrin (Schneider et Shiffrin, 1977; Shiffrin et Schneider, 1977) distinguant et caractérisant les processus automatiques et les processus contrôlés dans le traitement de l'information. Comme nous l'avons abordé dans le chapitre 1.1.1.2., ces processus présentent des caractéristiques différentes résumées dans le tableau 1.
Une telle distinction entre deux processus est encore actuellement étudiée par beaucoup d'auteurs dans différentes modalités. Les travaux de Posner présentés ci-dessus suggèrent également une dissociation entre processus endogènes et processus exogènes.
1.3.4. Le modèle de Laberge
Dans la perspective des travaux sur l'attention visuelle, Laberge (1975; 1995) a proposé un modèle portant, notamment, sur l'automatisation et la sélection attentionnelle. Les différentes informations d'un objet (par exemple les différents segments de la lettre 'b') convergeraient vers un noeud mnésique caractérisant l'encodage de l'objet (de la lettre 'b'). En fonction de l'utilisation du stimulus, un 'centre d'attention' pourrait venir ensuite activer ce noeud. L'attention agirait donc à un niveau assez tardif sur les noeuds mnésiques. Les signaux nerveux correspondant à l'objet focalisé bénéficieraient ainsi d'un rehaussement permettant de les 'faire sortir' du bruit neuronal général. Ce rehaussement correspondrait à une 'fenêtre attentionnelle' pouvant être brève (environ 100 ms) ou nettement plus longue (quelques secondes). Le premier type de fenêtre correspondrait à une focalisation de l'attention alors que le second type permettrait un processus d'attention préparatoire. Selon Laberge, la mise en jeu de l'attention n'est pas liée aux processus perceptifs, mais à la décision d'utiliser ou non un codage sensoriel réalisé automatiquement (au niveau du noeud).
En se basant sur différents travaux étudiant les structures cérébrales impliquées dans l'attention visuelle (dont certains ont été présentés ci-dessus), Laberge (1995) intègre différentes structures cérébrales dans les processus attentionnels (l'aire V4, le cortex pariétal postérieur, le cortex inféro-temporal ainsi que plusieurs régions du cortex préfrontal). L'orientation de l'attention serait assurée par les tubercules quadrijumeaux supérieurs et par le pulvinar. Ce modèle permet de rendre compte d'une grande variété de processus attentionnels, dont ceux d'orientation de l'attention visuo-spatiale (comme le modèle de Posner). Cependant, ce modèle ne prend pas en compte les différences fonctionnelles qui semblent exister entre l'hémisphère gauche et l'hémisphère droit comme le témoignent, notamment, les déficits observés dans l'héminégligence. En revanche, le modèle de Mesulam (1981) se base sur ces différences hémisphériques de l'attention (chapitre 1.3.5.).
1.3.4. Le modèle de Posner
Posner et ses collaborateurs (pour une revue voir Posner et Raichle, 1994) ont étudié les processus d'orientation visuo-spatiale de l'attention dans des tâches que nous avons présentées dans le chapitre 1.1.1.1. Ces études ont mis en évidence d'une part deux types d'orientation de l'attention: l'orientation exogène et l'orientation endogène et d'autre part une série d'opérations mentales impliquées dans l'orientation de l'attention. En se basant sur des études effectuées sur des animaux (lésions restreintes de certaines zones cérébrales) ainsi que sur les observations de patients cérébro-lésés (présentant des difficultés spécifiques selon l'emplacement de leurs lésions cérébrales), ces auteurs ont spécifié les aires cérébrales et les réseaux sous-jacents aux processus attentionnels observés dans leurs études comportementales. Trois zones cérébrales seraient impliquées dans ces processus: le lobe pariétal postérieur, le colliculus supérieur et le pulvinar (Fig. 2). A partir de ces trois zones, ces auteurs (Posner et Petersen, 1990; Posner et Rothbart, 1991; Posner, 1994) suggèrent la présence de trois réseaux attentionnels sous-jacents à ces différentes opérations attentionnelles: un réseau attentionnel postérieur, un réseau attentionnel antérieur et un réseau de vigilance (Fig. 9).

Ces réseaux auraient donc des fonctions différentes (Posner et Petersen, 1990). Le réseau postérieur serait responsable de l'orientation de l'attention vers des stimuli pertinents. Le cortex pariétal serait impliqué dans les processus de désengagement de l'attention d'un endroit donné. Le colliculus supérieur agirait au niveau du mouvement de l'attention vers un nouvel emplacement et le pulvinar permettrait l'engagement de l'attention sur la nouvelle cible (voir également Fig. 5). Le réseau attentionnel antérieur serait impliqué dans la détection. Il serait constitué du cortex cingulaire et de l'aire motrice supplémentaire dont les rôles dans la détection d'objets et la sélection de la réponse appropriée ont été souvent suggérés (voir chapitres 1.1.1. et 1.1.2. ci-dessus). Le troisième réseau impliquerait le locus coeruleus et les connexions de cette structure avec le lobe pariétal et le colliculus. Les connexions noradrénergiques du locus coeruleus au cortex seraient cruciales dans le maintien de la vigilance (Harley, 1987). Des études en imagerie fonctionnelle ont mis en évidence une augmentation d'activité dans la région du cortex frontal latéral droit dans des tâches de vigilance (Cohen et al., 1988; Pardo et al., 1991). Ce troisième réseau semble donc être latéralisé spécifiquement dans l'hémisphère droit.
Ces trois réseaux seraient indépendants des systèmes sensoriels, moteur ou des autres réseaux cognitifs (comme le langage, la mémoire, etc.). Ils seraient liés les uns aux autres par plusieurs connexions. Par exemple, chez le singe, Goldman-Rakic (1988) a mis en évidence des connexions entre le gyrus cingulaire et le lobe pariétal postérieur qui témoigneraient de l'existence d'une relation entre le système antérieur et le système postérieur. Ces réseaux bien individualisés peuvent expliquer que des lésions focalisées dans certaines de ces zones produisent des troubles purement attentionnels. Ce modèle permet également de différencier les processus attentionnels de l'expérience consciente. L'orientation de l'attention serait ainsi sous la dépendance de l'intégrité des systèmes attentionnels postérieurs, alors que l'expérience consciente impliquerait l'intégrité des réseaux attentionnels antérieurs (Posner et Rothbart, 1991). Les mêmes cérébrales prises en compte dans le modèle de Posner sont les mêmes que celles considérées dans le modèle de Mesulam (1981, 1990). L'organisation des régions ainsi que les fonctions attribuées à ces régions sont cependant différentes entre ces deux modèles.
1.3.5. Le modèle de Mesulam
Le modèle développé par Mesulam (1981; 1990) se base principalement sur les observations de patients cérébro-lésés et sur les études neuroanatomiques des primates non humains. Son modèle propose l'existence d'un réseau prenant en compte le cortex pariétal supérieur, le cortex cingulaire et la région appelée 'frontal eye field' (FEF, correspondant à l'aire 8 dans la nomenclature de Brodmann). Ces trois régions seraient toutes sous l'influence du système réticulé activateur ascendant qui fournirait un niveau d'éveil correct (Fig. 10). Selon ce modèle, un système de coordonnées spatiales serait présent dans chacune de ces régions. La région pariétale contiendrait une carte perceptive interne du monde extérieur. Le gyrus cingulaire régulerait la distribution spatiale de l'attention alors que la région frontale coordonnerait les programmes moteurs d'exploration, d'atteinte et de fixation.
Ces différentes régions sont interconnectées les unes aux autres, ainsi qu'à des structures sous-corticales. Ces dernières sont le colliculus supérieur (connecté au FEF et au cortex pariétal notamment), le pulvinar et le striatum (connectés à un grand nombre de régions corticales, Fries, 1984). Finalement, les trois régions principales de ce modèle sont également connectées à des zones corticales comme le cortex inféro-temporal et le cortex orbito-frontal. Cependant, les zones impliquées spécifiquement dans l'attention sont les cortex pariétal, cingulaire et frontal.
1.3.6. Conclusion
Les différents modèles abordés dans la première partie de ce chapitre (chapitre 1.3) prennent en compte les différents processus impliqués dans l'attention. Broadbent ainsi que Treisman ont précisé le rôle limitatif de l'attention qui permet la sélection des informations pertinentes. Dans leurs travaux sur l'attention visuelle, Schneider et Shiffrin ont mis en évidence la présence de deux processus distincts: les processus automatiques et contrôlés. Ils ont proposé une dissociation entre les notions d'automatique et de contrôlé qui a été reprise dans plusieurs modèles de l'attention, dont celui de Posner (à travers les fonctions respectives de ses trois réseaux).
Nous allons aborder dans le prochain chapitre le syndrome de l'héminégligence, caractérisé par des déficits attentionnels qui seraient consécutifs à des lésions situées principalement dans certaines régions prises en compte dans ces modèles. Comme nous l'avons décrit dans ce premier chapitre, la majorité des études sur l'attention ont porté sur une seule modalité. Bien que les processus attentionnels auditifs et visuels semblent être assez similaires, l'existence de zones cérébrales impliquées dans l'attention communes aux deux modalités est encore en question. Nos travaux ont porté sur les processus attentionnels automatiques et contrôlés dans les modalités auditive et visuelle. Ils avaient pour but de spécifier les relations entre ces processus dans ces deux modalités en étudiant des patients cérébro-lésés présentant un syndrome d'héminégligence.